Порфирии. Зачет биохимия I
Серповидно-клеточная анемия
HbS – гемоглобин серповидно-клеточной анемии. При этом нарушении в ДНК в результате точковой мутации триплет ЦТТ заменен на триплет ЦАТ , что влечет за собой включение в 6-м положении β-цепи вместо глутамата аминокислоты валина . Изменение свойств β-цепи влечет изменение свойств всей молекулы и формирование на поверхности гемоглобина "липкого" участка. При дезоксигенации гемоглобина участок "раскрывается" и связывает одну молекулу дезоксигемоглобина S с другими подобными. Результатом является полимеризация гемоглобиновых молекул и образование крупных белковых тяжей, вызывающих деформацию эритроцитов и, при прохождении ими капилляров, гемолиз.
Схема отличия гемоглобина S от гемоглобина А и его полимеризация
Нарушение синтеза гемоглобина
Порфирии
Порфирии – это группа гетерогенных наследственных заболеваний, возникающих в результате нарушения синтеза гема и повышения содержания порфиринов и их предшественников в организме. Выделяют наследственные и приобретенные формы порфирии.
Приобретенные формы порфирий носят токсический характер и вызываются действием гексахлорбензола, солей свинца и других тяжелых металлов (ингибирование порфобилиногенсинтазы, феррохелатазы и др.), лекарственными препаратами (антигрибковый антибиотик гризеофульфин).
При наследственных формах дефект фермента имеется во всех клетках организма, но проявляется только в одном типе клеток. Можно выделить две большие группы порфирий:
1. Печеночные – группа заболеваний с аутосомно-доминантными нарушениями ферментов различных этапов синтеза протопорфирина IX .
Наиболее ярким заболеванием этой группы является перемежающаяся острая порфирия , при которой у гетерозигот активность уропорфириноген-I-синтазы снижена на 50%. В результате больные экскретируют с мочой большие количества порфобилиногена и аминолевулиновой кислоты. На свету порфобилиноген окисляется в окрашенные соединения и моча темнеет. Симптомами заболевания являются острые боли в животе, нервно-психические расстройства (полиневриты, тетрапарез, галлюцинации), сердечно-сосудистые нарушения.
Заболевание проявляется после достижения половой зрелости из-за повышенной потребности гепатоцитов в цитохроме Р450 для обезвреживания половых стероидов, обострение состояния также часто бывает после приема лекарственных препаратов, метаболизм которых требует участия цитохрома Р450 . Снижение концентрации гема, используемого для синтеза цитохрома Р450 , активирует аминолевулинат синтазу .
2. Эритропоэтические – аутосомно-рецессивные нарушения некоторых ферментов синтеза протопорфирина IX в эритроидных клетках. При этом при синтезе уропорфириногенов
Гем является простетической группой многих белков: гемоглобина, миоглобина, цитохромов митохондриальной ЦПЭ, цитохрома Р 450 , участвующего в микросомальном окислении. Ферменты каталаза, пероксидаза, цитохромоксидаза содержат гем в качестве кофермента.
Все клетки организма имеют гемсодержащие белки, поэтому синтез гема идёт во всех клетках, за исключением эритроцитов, не имеющих, как известно, белоксинтезирующей системы.
При распаде гема в клетках РЭС образуется жёлчный пигмент билирубин. Дальнейший катаболизм билирубина в печени, кишечнике и почках приводит к образованию конечных продуктов распада гема стеркобилина и уробилина, содержащихся, соответственно, в кале и моче. Железо, освобождающееся при распаде гема, снова используется для синтеза железосодержащих белков.
I. СТРОЕНИЕ И БИОСИНТЕЗ ГЕМА а. строение гема
Гем состоит из иона двухвалентного железа и порфирина (рис. 13-1). В основе структуры пор-фиринов находится порфин. Порфин представляет собой четыре пиррольных кольца, связанных между собой метеновыми мостиками (рис. 13-1). В зависимости от структуры заместителей в кольцах пирролов различают несколько типов порфиринов: протопорфирины, этиопорфи-рины, мезопорфирины и копропорфирины. Протопорфирины - предшественники всех других типов порфиринов.
Гемы разных белков могут содержать разные типы порфиринов (см. раздел 6). В геме гемоглобина находится протопорфирин IX, который имеет 4 метильных, 2 винильных радикала и 2 остатка пропионовой кислоты. Железо в геме находится в восстановленном состоянии (Fе +2) и связано двумя ковалентными и двумя координационными связями с атомами азота пиррольных колец. При окислении железа гем превращается
в гематин (Fe 3 +). Hаибольшее количество гема содержат эритроциты, заполненные гемоглобином, мышечные клетки, имеющие миоглобин, и клетки печени из-за высокого содержания в них цитохрома Р 450 .
б. биосинтез гема
Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени (рис. 13-2). В костном мозге гем необходим для синтеза гемоглобина в ретикулоцитах, в гепато-цитах - для образования цитохрома Р 450 .
Первая реакция синтеза гема - образование 5-аминолевулиновой кислоты из глицина и сукцинил-КоА (рис. 13-3) идёт в матриксе митохондрий, где в ЦТК образуется один из субстратов этой реакции - сукцинил-КоА. Эту реакцию катализирует пиридоксальзависимый фермент 5-аминолевулинатсинтаза.
Из митохондрий 5-аминолевулиновая кислота поступает в цитоплазму. В цитоплазме проходят промежуточные этапы синтеза гема: соединение 2 молекул 5-аминолевулиновой кислоты в молекулу порфобилиногена (рис. 13-4), дезаминирование порфобилиногена с образованием гидроксиметилбилана, ферментативное превращение гидроксиметилбилана в молекулу уропорфобилиногена III, декар-боксилирование последнего с образованием копропорфириногена III. Гидроксиметилбилан может также неферментативно превращаться в уропорфириноген I, который декарбоксилиру-ется в копропорфириноген I. Из цитоплазмы копропорфириноген III опять поступает в митохондрии, где проходят заключительные реакции синтеза гема. В результате двух последовательных окислительных реакций коп-ропорфириноген III превращается в протопор-фириноген IX, а протопорфириноген IX - в протопорфирин IX. Фермент феррохелатаза, присоединяя к протопорфирину IX двухвалентное железо, превращает его в гем (рис. 13-2). Источником железа для синтеза гема служит депонирующий железо белок ферритин. Син-
Рис. 13-1. Строение порфина (А), протопорфирина IX (Б) и гема гемоглобина (В). Порфин - циклическая структура, состоящая из четырёх пиррольных колец, связанных между собой метеновыми мостиками. Прото-порфирин IX имеет четыре метильных, два винильных радикала и два остатка пропионовой кислоты. В геме гемоглобина Fe 2+ образует две ковалентные и две координационные связи с атомами азота пиррольных колец протопорфирина IX.
тезированный гем, соединяясь с α- и β-по-липепептидными цепями глобина, образует гемоглобин. Гем регулирует синтез глобина: при снижении скорости синтеза гема синтез глобина в ретикулоцитах тормозится.
В. РЕГУЛЯЦИЯ БИОСИНТЕЗА ГЕМА
Регуляторную реакцию синтеза гема катализирует пиридоксальзависимый фермент 5-аминолевулинатсинтаза. Скорость реакции регулируется аллостерически и на уровне трансляции фермента.
Аллостерическим ингибитором и корепрессо-ром синтеза 5-аминолевулинатсинтазы является гем (рис. 13-5).
В ретикулоцитах синтез этого фермента на этапе трансляции регулирует железо. На участке инициации мРHК, кодирующей фермент, имеется последовательность нуклеотидов, образующая шпилечную петлю, которая называется железочувствительным элементом (от англ. iron-responsive element, IRE) (рис. 13-6).
При высоких концентрациях железа в клетках оно образует комплекс с остатками цисте-ина регуляторного железосвязывающего белка. Взаимодействие железа с регуляторным желе-зосвязывающим белком вызывает снижение сродства этого белка к IRE-элементу мРHК, кодирующей 5-аминолевулинатсинтазу, и продолжение трансляции (рис. 13-6, А). При низких концентрациях железа железосвязывающий

Рис. 13-2. Синтез гема. Цифрами на схеме указаны ферменты: 1 - 5-аминолевулинатсинтаза; 2 - 5-аминоле-вулинатдегидратаза; 3 - порфобилиногендезаминаза; 4 - уропорфириноген III косинтаза; 5 - уропорфирино-гендекарбоксилаза; 6 - копропорфириноген III оксидаза; 7 - протопорфириногеноксидаза; 8 - феррохелатаза. Буквами обозначены заместители в пиррольных кольцах: М - метил, В - винил, П - остатки пропионовой кислоты, А - ацетил, ПФ - пиридоксальфосфат. Донором железа служит депонирующий железо в клетках белок ферритин.

Рис. 13-3. Реакция образования 5-аминолевулиновой кислоты.
белок присоединяется к железочувствительному элементу, находящемуся на 5"-нетранслируемом конце мРНК, и трансляция 5-аминолевулинат-синтазы тормозится (рис. 13-6, Б).
5-Аминолевулинатдегидратаза также аллостери-чески ингибируется гемом, но так как активность этого фермента почти в 80 раз превышает активность 5-аминолевулинатсинтазы, то это не имеет большого физиологического значения.

Рис. 13-4. Реакция образования порфобилиногена.

Рис. 13-5. Регуляция синтеза гема и гемоглобина. Гем по принципу отрицательной обратной связи ингибирует 5-аминолевулинатсинтазу и 5-аминолевулинатдегидратазу и является индуктором трансляции α- и β -цепей гемо глобина.

Рис. 13-6. Регуляция синтеза аминолевулинатсинтазы. А - при высокой концентрации железа в ретикулоцитах оно присоединяется к железосвязывающему белку и снижает сродство этого белка к железочувствительному элементу (IRE) матричной РНК, кодирующей 5-аминолевулинатсинтазу. Белковые факторы инициации трансляции связываются с мРНК и инициируют трансляцию 5-аминолевулинатсинтазы. Б - при низком содержании железа в ретикулоцитах железосвязывающий белок обладает высоким сродством к IRE и взаимодействует с ним. Белковые факторы инициации трансляции не могут присоединиться к мРНК, и трансляция прекращается.
Дефицит пиридоксальфосфата и лекарственные препараты, которые являются его структурными аналогами, снижают активность 5-амино-левулинатсинтазы.
Г. НАРУШЕНИЯ БИОСИНТЕЗА ГЕМА. ПОРФИРИИ
Hаследственные и приобретённые нарушения синтеза гема, сопровождающиеся повышением содержания порфириногенов, а также продуктов их окисления в тканях и крови и появлением их в моче, называют порфириями («порфирин» в переводе с греч. означает пурпурный). Моча больных имеет красный цвет.
Hаследственные порфирии обусловлены генетическими дефектами ферментов, участвующих
в синтезе гема, за исключением 5-аминолевули-натсинтазы. При этих заболеваниях отмечают снижение образования гема. Поскольку гем - ал-лостерический ингибитор 5-аминолевулинатсин-тазы, то активность этого фермента повышается, и это приводит к накоплению промежуточных продуктов синтеза гема - 5-аминолевулиновой кислоты и порфириногенов.
В зависимости от основной локализации патологического процесса различают печёночные и эритропоэтические наследственные порфирии. Эритропоэтические порфирии сопровождаются накоплением порфиринов в нормобластах и эритроцитах, а печёночные - в гепатоцитах.
При тяжёлых формах порфирий наблюдают нейропсихические расстройства, нарушения функций РЭС, повреждения кожи. Порфири-ногены не окрашены и не флуоресцируют, но на свету они легко превращаются в порфирины. Последние проявляют интенсивную красную флуоресценцию в ультрафиолетовых лучах. В коже на солнце в результате взаимодействия с порфиринами кислород переходит в синглетное состояние. Синглетный кислород вызывает ускорение ПОЛ клеточных мембран и разрушение клеток, поэтому порфирии часто сопровождаются фотосенсибилизацией и изъязвлением открытых участков кожи. Нейропсихические расстройства при порфириях связаны с тем, что 5-аминолевулинат и порфириногены являются нейротоксинами.
Иногда при лёгких формах наследственных порфирий заболевание может протекать бессимптомно, но приём лекарств, являющихся индукторами синтеза 5-аминолевулинатсинтазы, может вызвать обострение болезни. Индукторами синтеза 5-аминолевулинатсинтазы являются такие известные лекарства, как сульфанил-ами-ды, барбитураты, диклофенак, вольтарен, стероиды, гестагены. В некоторых случаях симптомы болезни не проявляются до периода полового созревания, когда повышение образования β-стероидов вызывает индукцию синтеза 5-ами-нолевулинатсинтазы. Порфирии наблюдают и при отравлениях солями свинца, так как свинец ингибирует 5-аминолевулинатдегидратазу и феррохелатазу. Некоторые галогенсодержащие гербициды и инсектициды являются индукторами синтеза 5-аминолевулинатсинтазы, поэтому попадание их в организм сопровождается симптомами порфирии.
II. ОБМЕН ЖЕЛЕЗА
В организме взрослого человека содержится 3-4 г железа, из которых только около 3,5 мг
находится в плазме крови. Гемоглобин имеет примерно 68% железа всего организма, ферри-тин - 27%, миоглобин - 4%, трансферрин - 0,1%, На долю всех содержащих железо ферментов приходится всего 0,6% железа, имеющегося в организме. Источниками железа при биосинтезе железосодержащих белков служат железо пищи и железо, освобождающееся при постоянном распаде эритроцитов в клетках печени и селезёнки.
В нейтральной или щелочной среде железо находится в окисленном состоянии - Fe 3+ , образуя крупные, легко агрегирующие комплексы с ОН - , другими анионами и водой. При низких значениях рН железо восстанавливается и легко диссоциирует. Процесс восстановления и окисления железа обеспечивает его перераспределение между макромолекулами в организме. Ионы железа обладают высоким сродством ко многим соединениям и образуют с ними хелатные комплексы, изменяя свойства и функции этих соединений, поэтому транспорт и депонирование железа в организме осуществляют особые белки. В клетках железо депонирует белок ферритин, в крови его транспортирует белок трансферрин.
а. всасывание железа в кишечнике
В пище железо в основном находится в окисленном состоянии (Fe 3+) и входит в состав белков или солей органических кислот. Освобождению железа из солей органических кислот способствует кислая среда желудочного сока. Наибольшее количество железа всасывается в двенадцатиперстной кишке. Аскорбиновая кислота, содержащаяся в пище, восстанавливает железо и улучшает его всасывание, так как в клетки слизистой оболочки кишечника поступает только Fe 2+. В суточном количестве пищи обычно содержится 15-20 мг железа, а всасывается только около 10% этого количества. Организм взрослого человека теряет около 1 мг железа в сутки.
Количество железа, которое всасывается в клетки слизистой оболочки кишечника, как правило, превышает потребности организма. Поступление железа из энтероцитов в кровь зависит от скорости синтеза в них белка апоферритина. Апоферритин «улавливает» железо в энтероцитах и превращается в ферритин, который остаётся в энтероцитах. Таким способом снижается поступ-
ление железа в капилляры крови из клеток кишечника. Когда потребность в железе невелика, скорость синтеза апоферритина повышается (см. ниже «Регуляция поступления железа в клетки»). Постоянное слущивание клеток слизистой оболочки в просвет кишечника освобождает организм от излишков железа. При недостатке железа в организме апоферритин в энтероцитах почти не синтезируется. Железо, поступающее из энтероцитов в кровь, транспортирует белок плазмы крови трансферрин (рис. 13-7).
Б. ТРАНСПОРТ ЖЕЛЕЗА В ПЛАЗМЕ КРОВИ И ЕГО ПОСТУПЛЕНИЕ В КЛЕТКИ
В плазме крови железо транспортирует белок трансферрин. Трансферрин - гликопротеин, который синтезируется в печени и связывает только окисленное железо (Fe 3+). Поступающее в кровь железо окисляет фермент ферроксидаза, известный как медьсодержащий белок плазмы крови церулоплазмин. Одна молекула трансфер-рина может связать один или два иона Fe 3+ , но одновременно с анионом СО 3 2- с образованием комплекса трансферрин-2 (Fе 3+ -СО 3 2-). В норме трансферрин крови насыщен железом приблизительно на 33%.
Трансферрин взаимодействует со специфическими мембранными рецепторами клеток.
В результате этого взаимодействия в цитозоле клетки образуется комплекс Са 2+ -кальмодулин-ПКС, который фосфорилирует рецептор транс-феррина и вызывает образование эндосомы. АТФ-зависимый протонный насос, находящийся в мембране эндосомы, создаёт кислую среду внутри эндосомы. В кислой среде эндосомы железо освобождается из трансферрина. После этого комплекс рецептор-апотрансферрин возвращается на поверхность плазматической мембраны клетки. При нейтральном значении рН внеклеточной жидкости апотрансферрин изменяет свою конформацию, отделяется от рецептора, выходит в плазму крови и становится способным вновь связывать ионы железа и включаться в новый цикл его транспорта в клетку. Железо в клетке используется для синтеза железосодержащих белков или депонируется в белке ферритине.
Ферритин - олигомерный белок с молекулярной массой 500 кД. Он состоит из тяжёлых (21 кД) и лёгких (19 кД) полипептидных цепей, составляющих 24 протомера. Разный набор протомеров в олигомере ферритина определяет образование нескольких изоформ этого белка в разных тканях. Ферритин представляет собой полую сферу, внутри которой может содержаться до 4500 ионов трёхвалентного железа, но обычно содержится менее 3000. Тяжёлые цепи

Рис. 13-7. Поступление экзогенного железа в ткани. В полости кишечника железо освобождается из белков и солей органических кислот пищи. Усвоению железа способствует аскорбиновая кислота, восстанавливающая железо. В клетках слизистой оболочки кишечника избыток поступившего железа соединяется с белком апофер-ритином с образованием ферритина, при этом ферритин окисляет Fe2+ в Fe3+. Поступление железа из клеток слизистой оболочки кишечника в кровь сопровождается окислением железа ферментом сыворотки крови фер-роксидазой. В крови Fe3+ транспортирует белок сыворотки крови трансферрин. В тканях Fe2+ используется для синтеза железосодержащих белков или депонируется в ферритине.
ферритина окисляют Fe 2+ в Fe 3+ . Железо в виде гидроксидфосфата находится в центре сферы, оболочка которой образована белковой частью молекулы. Оно поступает внутрь и освобождается наружу через каналы, пронизывающие белковую оболочку апоферритина, но железо может откладываться и в белковой части молекулы ферритина. Ферритин содержится почти во всех тканях, но в наибольшем количестве в печени, селезёнке и костном мозге. Незначительная часть ферритина экскретируется из тканей в плазму крови. Поскольку поступление ферри-тина в кровь пропорционально его содержанию в тканях, то концентрация ферритина в крови - важный диагностический показатель запасов железа в организме при железодефицитной анемии. Метаболизм железа в организме представлен на рис. 13-8.
В. РЕГУЛЯЦИЯ ПОСТУПЛЕНИЯ ЖЕЛЕЗА В КЛЕТКИ
Содержание железа в клетках определяется соотношением скоростей его поступления, использования и депонирования и контролируется двумя молекулярными механизмами. Скорость поступления железа в неэритроидные клетки зависит от количества белков-рецепторов трансферрина в их мембране. Избыток железа в клетках депонирует ферритин. Синтез апофер-ритина и рецепторов трансферрина регулируется на уровне трансляции этих белков и зависит от содержания железа в клетке.
На нетранслируемом 3"-конце мРНК рецептора трансферрина и на нетранслируемом 5"-конце мРНК апоферритина имеются шпилечные петли - железочувствительные элементы IRE (рис. 13-9 и 13-10). Причём мРНК рецептора трансфер-

Рис. 13-8. Метаболизм железа в организме.

Рис. 13-9. Регуляция синтеза апоферритина. А - при снижении содержания железа в клетке железосвязывающий белок обладает высоким сродством к IRE и взаимодействует с ним. Это препятствует присоединению белковых факторов инициации трансляции к мРНК, кодирующей апоферритин, и синтез апоферритина прекращается; Б - при повышении содержания железа в клетке оно взаимодействует с железосвязывающим белком, в результате чего снижается сродство этого белка к IRE. Белковые факторы инициации трансляции присоединяются к мРНК, кодирующей апоферритин, и инициируют трансляцию апоферритина.
рина имеет 5 петель, а мРНК апоферритина - только 1.
Эти участки мРНК могут взаимодействовать с регуляторным IRE-связывающим белком. При низких концентрациях железа в клетке IRE-связывающий белок соединяется с IRE мРНК апоферритина и препятствует присоединению белковых факторов инициации трансляции (рис. 13-9, А). В результате этого снижаются скорость трансляции апоферри-тина и его содержание в клетке. Вместе с тем при низких концентрациях железа в клетке IRE-связывающий белок связывается с желе-зочувствительным элементом мРНК рецептора трансферрина и предотвращает её разрушение ферментом РНК-азой (рис. 13-10, А). Это вызывает увеличение количества рецепторов
трансферрина и ускорение поступления железа в клетки.
При повышении содержания железа в клетке в результате его взаимодействия с IRE-связывающим белком происходит окисление SH-групп активного центра этого белка и снижение сродства к железочувствительным элементам мРНК. Это приводит к двум последствиям:
Во-первых, ускоряется трансляция апоферритина (рис. 13-9, Б);
Во-вторых, IRE-связывающий белок освобождает шпилечные петли мРНК рецептора трансферрина, и она разрушается ферментом РНК-азой, в результате снижается скорость синтеза рецепторов трансферрина (рис. 1310, Б). Ускорение синтеза апоферритина и торможение синтеза рецепторов транс-

Рис. 13-10. Регуляция синтеза рецептора трансферрина. А - при низком содержании железа в клетке железочувствительный белок обладает высоким сродством к IRE мРНК, кодирующей белок-рецептор транс-феррина. Присоединение железосвязывающего белка к IRE мРНК предотвращает её разрушение РНК-азой и синтез белка-рецептора трансферрина продолжается; Б - При высоком содержании железа в клетке сродство железосвязывающего белка к IRE снижается, и мРНК становится доступной для действия РНК-азы, которая её гидролизует. Разрушение мРНК ведёт к снижению синтеза белка-рецептора трансферрина.
феррина вызывают снижение содержания
железа в клетке. В целом эти механизмы регулируют содержание железа в клетках и его использование для синтеза железосодержащих белков.
Г. НАРУШЕНИЯ МЕТАБОЛИЗМА ЖЕЛЕЗА
Железодефицитная анемия может наблюдаться при повторяющихся кровотечениях, беременности, частых родах, язвах и опухолях
ЖКТ, после операций на ЖКТ. При желе-зодефицитной анемии уменьшается размер эритроцитов и их пигментация (гипохромные эритроциты малых размеров). В эритроцитах уменьшается содержание гемоглобина, понижается насыщение железом трансферрина, а в тканях снижается концентрация ферритина. Причина этих изменений - недостаток железа в организме, вследствие чего снижается синтез гема и ферритина в неэритроидных тканях и гемоглобина в эритроидных клетках.
Гемохроматоз. Когда количество железа в клетках превышает объём ферритинового депо, железо откладывается в белковой части молекулы ферритина. В результате образования таких аморфных отложений избыточного железа ферритин превращается в гемосидерин. Гемо-сидерин плохо растворим в воде и содержит до 37% железа. Накопление гранул гемосидерина в печени, поджелудочной железе, селезёнке приводит к повреждению этих органов - гемохроматозу. Гемохроматоз может быть обусловлен наследственным увеличением всасывания железа в кишечнике, при этом содержание железа в организме больных может достигать 100 г. Это заболевание наследуется по ауто-сомно-рецессивному типу, причём около 0,5% европеоидов гомозиготны по гену гемохрома-тоза. Накопление гемосидерина в поджелудочной железе приводит к разрушению β-клеток островков Лангерханса и, как следствие этого, к сахарному диабету. Отложение гемосидерина в гепатоцитах вызывает цирроз печени, а в ми-окардиоцитах - сердечную недостаточность. Больных наследственным гемохроматозом лечат регулярными кровопусканиями, еженедельно или один раз в месяц в зависимости от тяжести состояния больного. К гемохроматозу могут привести частые переливания крови, в этих случаях больных лечат препаратами, связывающими железо.
III. КАТАБОЛИЗМ ГЕМОГЛОБИНА
Эритроциты имеют короткое время жизни (примерно 120 дней). При физиологических условиях в организме взрослого человека разрушается около 1-2х10 и эритроцитов в сутки. Их катаболизм происходит главным образом в ретикулоэндотелиальных клетках селезёнки, лимфатических узлов, костного мозга и печени. При старении эритроцитов снижается содержание сиаловых кислот в составе гликопротеинов плазматической мембраны. Изменённые углеводные компоненты гликопротеинов мембран эритроцитов связываются рецепторами клеток РЭС, и эритроциты «погружаются» в них эн-доцитозом. Распад эритроцитов в этих клетках начинается с распада гемоглобина на гем и глобин и последующего гидролиза ферментами лизосом белковой части гемоглобина.
А. КАТАБОЛИЗМ ГЕМА
Первая реакция катаболизма гема происходит при участии NADРН-зависимого ферментативного комплекса гемоксигеназы. Ферментная система локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, - таким образом, раскрывается структура кольца (рис. 13-11). В ходе реакции образуются линейный тетрапир-рол - биливердин (пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к гему.
Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADРН-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, но также при катаболизме других гемсодержащих белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека - примерно 250-350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени.
Б. МЕТАБОЛИЗМ БИЛИРУБИНА
Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), плохо растворим в воде, по крови транспортируется в комплексе с белком плазмы крови альбумином. Эту форму билирубина называют неконъюгированным билирубином. Каждая молекула альбумина связывает 2 (или даже 3) молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге рН крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за

Рис. 13-11. Распад гема. М - (-СН 3) - метильная группа; В - (-СН=СН 2) - винильная группа; П - (-СН 2 -СН 2 -СООН) - остаток пропионовой кислоты. В ходе реакции одна метильная группа превращается в окись углерода и, таким образом, раскрывается структура кольца. Образованный биливердин под действием биливердинредуктазы превращается в билирубин.
высокоаффинный, имеющий высокое сродство центр альбумина.
Поглощение билирубина паренхиматозными клетками печени
Комплекс «альбумин-билирубин», доставляемый с током крови в печень, на поверх-
ности плазматической мембраны гепатоцита диссоциирует. Высвобожденный билирубин образует временный комплекс с липидами плазматической мембраны. Облегчённая диффузия билирубина в гепатоциты осуществляется двумя типами белков-переносчиков: лигандина (он транспортирует основное количество били-
рубина) и протеина Z. Активность поглощения билирубина гепатоцитом зависит от скорости его метаболизма в клетке.
Лигандин и протеин Z обнаружены также в клетках почек и кишечника, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детокси-кации в этом органе.
Конъюгация билирубина в гладком ЭР
В гладком ЭР гепатоцитов к билирубину присоединяются остатки глюкуроновой кислоты - реакции конъюгации. Билирубин имеет 2 карбоксильные группы, поэтому может соединяться с 2 молекулами глюкуроновой кислоты, образуя хорошо растворимый в воде конъюгат - ди-глюкуронид билирубина (конъюгированный, или прямой, билирубин) (рис. 13-12).
Донором глюкуроновой кислоты служит УДФ-глюкуронат. Специфические ферменты, УДФ-глюкуронилтрансферазы (уридиндифосфоглю-куронилтрансферазы) катализируют образование моно- и диглюкуронидов билирубина (рис. 13-13). Индукторами синтеза УДФ-глюкуронилтрансфераз служат некоторые лекарственные препараты, например, фенобарбитал (см. раздел 12).
Секреция билирубина в жёлчь
Секреция конъюгированного билирубина в жёлчь идёт по механизму активного транспорта, т.е. против градиента концентрации. Активный транспорт является, вероятно, скорость-лими-тирующей стадией всего процесса метаболизма билирубина в печени. В норме диглюкуронид

Рис. 13-12. Структура билирубиндиглюкуронида (конъюгированный, «прямой» билирубин). Глюку-роновая кислота присоединяется эфирной связью к двум остаткам пропионовой кислоты с образованием ацилглюкуронида.
билирубина - главная форма экскреции билирубина в жёлчь, однако не исключается присутствие небольшого количества моно-глюкуронида. Транспорт конъюгированного билирубина из печени в жёлчь активируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, можно сказать, что скорость конъюгации билирубина и активный транспорт билирубинглюкуронида из гепатоцитов в жёлчь строго взаимосвязаны (рис. 13-14).
В. КАТАБОЛИЗМ БИЛИРУБИНДИГЛЮКУРОНИДА
В кишечнике поступившие билирубинглюку-рониды гидролизуются специфическими бактериальными ферментами β-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты. Освобо-

Рис. 13-13. Образование билирубиндиглюкуронида.

Рис. 13-14. Билирубин-уробилиногеновый цикл в печени. 1 - катаболизм Hb в ретикулоэндотелиальных клетках костного мозга, селезёнки, лимфатических узлов; 2 - образование транспортной формы комплекса билирубин-альбумин; 3 - поступление билирубина в печень; 4 - образование билирубинглюкуронидов;
5 - секреция билирубина в составе жёлчи в кишечник;
6 - катаболизм билирубина под действием кишечных бактерий; 7 - удаление уробилиногенов с калом; 8 - всасывание уробилиногенов в кровь; 9 - усвоение уробилиногенов печенью; 10 - поступление части уробилиногенов в кровь и выделение почками с мочой; 11 - небольшая часть уробилиногенов секретируется в жёлчь.
дившийся в ходе этой реакции билирубин под действием кишечной микрофлоры восстанавливается с образованием группы бесцветных тетрапиррольных соединений - уробилиногенов (рис. 13-15).
В подвздошной и толстой кишках небольшая часть уробилиногенов снова всасывается, попадает с кровью воротной вены в печень. Основная часть уробилиногена из печени в составе жёлчи выводится в кишечник и выделяется с
калом из организма, часть уробилиногена из печени поступает в кровь и удаляется с мочой в форме уробилина (рис. 13-14). В норме большая часть бесцветных уробилиногенов, образующихся в толстой кишке, под действием кишечной микрофлоры окисляется в прямой кишке до пигмента коричневого цвета уробилина и удаляется с калом. Цвет кала обусловлен присутствием уробилина.

Рис. 13-15. Структура некоторых жёлчных пигментов. Мезобилиноген - промежуточный продукт катаболизма билирубина в кишечнике.
iv. диагностическое значение
определения концентрации билирубина в биологических жидкостях человека
В настоящее время для определения содержания билирубина в сыворотке (плазме) крови используют предложенный в 1916 г. Ван дер Бергом метод определения билирубина в сыворотке крови, основанный на диазореакции.
В нормальном состоянии концентрация общего билирубина в плазме составляет 0,3-1 мг/дл (1,7-17 мкмоль/л), 75% от общего количества билирубина находится в неконъюгированной форме (непрямой билирубин). В клинике ко-нъюгированный билирубин называют прямым, потому что он водорастворим и может быстро взаимодействовать с диазореагентом, образуя соединение розового цвета, - это и есть прямая реакция Ван дер Берга. Неконъюгированньш билирубин гидрофобен, поэтому в плазме крови содержится в комплексе с альбумином и не реагирует с диазореактивом до тех пор, пока не добавлен органический растворитель, например этанол, который осаждает альбумин. Неконъ-югированный билирубин, взаимодействующий с азокрасителем только после осаждения белка, называют непрямым билирубином.
У больных с печёночно-клеточной патологией, сопровождающейся длительным повышением
концентрации конъюгированного билирубина, в крови обнаруживают третью форму плазменного билирубина, при котором билирубин ковалентно связан с альбумином, и поэтому его невозможно отделить обычным способом. В некоторых случаях до 90% общего содержания билирубина крови может находиться в этой форме.
А. ЖЕЛТУХИ
Причинами гипербилирубинемии могут быть увеличение образования билирубина, превышающее способность печени экскретировать его, или повреждение печени, приводящее к нарушению секреции билирубина в жёлчь в нормальных количествах. Гипербилирубинемию отмечают также при закупорке желчевыводящих протоков печени.
Во всех случаях содержание общего билирубина в крови повышается. При достижении определённой концентрации он диффундирует в ткани, окрашивая их в жёлтый цвет. Пожелтение тканей из-за отложения в них билирубина называют желтухой. Клинически желтуха может не проявляться до тех пор, пока концентрация билирубина в плазме крови не превысит верхний предел нормы более чем в 2,5 раза, т.е. не станет выше 50 мкмоль/л.
1. Гемолитическая (надпечёночная) желтуха
Известно, что способность печени образовывать глюкурониды и выделять их в жёлчь в 3- 4 раза превышает их образование в физиологических условиях. Гемолитическая (надпечёноч-ная) желтуха - результат интенсивного гемолиза эритроцитов. Она обусловлена чрезмерным образованием билирубина, превышающим
способность печени к его выведению. Гемолитическая желтуха развивается при исчерпании резервных возможностей печени. Основная причина надпечёночной желтухи - наследственные или приобретённые гемолитические анемии. При гемолитических анемиях, вызванных сепсисом, лучевой болезнью, дефицитом глюкозо-6-фосфатдегидрогеназы эритроцитов, талассемией, переливанием несовместимых групп крови, отравлением сульфаниламидами, количество освобождающегося из эритроцитов гемоглобина за сутки может доходить до 45 г
(при норме 6,25 г), что значительно увеличивает образование билирубина. Гипербилирубинемия у больных гемолитической желтухой обусловлена значительным повышением (103-171 мкмоль/л) в крови концентрации альбуминсвязанного неконъюгированного билирубина (непрямой билирубин). Образование в печени и поступление в кишечник больших количеств билиру-бинглюкуронидов (прямой билирубин) ведёт к усиленному образованию и выделению с калом и мочой уробилиногенов и более интенсивной их окраски (рис. 13-16).

Рис. 13-16. Билирубин-уробилиногеновый цикл при гемолитической желтухе. 1 - катаболизм Hb идёт с повышенной скоростью; 2 - в крови примерно в 10 раз повышена концентрация непрямого билирубина; 3 - альбумин высвобождается из комплекса билирубин-альбумин; 4 - активность реакции глюкуронирования возрастает, но она ниже, чем скорость образования билирубина; 5 - секреция билирубина в жёлчь повышена; 6, 7,10 - повышенное содержание уробилиногенов в кале и моче придаёт им более интенсивную окраску; уро-билиноген всасывается из кишечника в кровь (8) и попадает в печень по воротной вене (9).
Один из главных признаков гемолитической желтухи - повышение содержания в крови неконъюгированного (непрямого) билирубина. Это позволяет легко отличить её от механической (подпечёночной) и печёночно-клеточной (печёночной) желтух.
Неконъюгированный билирубин токсичен. Гидрофобный, липофильный неконъюгирован-ный билирубин, легко растворяясь в липидах мембраны и проникая вследствие этого в митохондрии, разобщает в них дыхание и окислительное фосфорилирование, нарушает синтез белка, поток ионов калия через мембрану клетки и органелл. Это отрицательно сказывается на состоянии ЦНС, вызывая у больных ряд характерных неврологических симптомов.
Желтуха новорождённых
Частная разновидность гемолитической желтухи новорождённых - «физиологическая желтуха», наблюдающаяся в первые дни жизни ребёнка. Причиной повышения концентрации непрямого билирубина в крови служит ускоренный гемолиз и недостаточность функции белков и ферментов печени, ответственных за поглощение, конъюгацию и секрецию прямого билирубина. У новорождённых не только снижена активность УДФ-глюкуронилтрансферазы, но и, по-видимому, недостаточно активно происходит синтез второго субстрата реакции конъюгации УДФ-глюкуроната.
Известно, что УДФ-глюкуронилтрансфера-за - индуцируемый фермент (см. раздел 12). Новорождённым с физиологической желтухой вводят лекарственный препарат фенобарбитал, индуцирующее действие которого было описано в разделе 12.
Одно из неприятных осложнений «физиологической желтухи» - билирубиновая энцефалопатия. Когда концентрация неконъюгирован-ного билирубина превышает 340 мкмоль/л, он проходит через гематоэнцефалический барьер головного мозга и вызывает его поражение.
2. Печёночно-клеточная (печёночная) желтуха
Печёночно-клеточная (печёночная) желтуха обусловлена повреждением гепатоцитов и жёлчных капилляров, например, при острых вирусных инфекциях, хроническом и токсических гепатитах.
Причина повышения концентрации билирубина в крови - поражение и некроз части печёночных клеток. Происходит задержка билирубина в печени, чему способствует резкое ослабление метаболических процессов в поражённых гепатоцитах, которые теряют способность нормально выполнять различные биохимические и физиологические функции, в частности переводить конъюгированный (прямой) билирубин из клеток в жёлчь против градиента концентрации. Для печёночно-клеточной желтухи характерно то, что вместо преобладающих в норме диглюкуронидов билирубина в поражённой печёночной клетке образуются главным образом моноглюкурониды
(рис. 13-17).
В результате деструкции печёночной паренхимы образующийся прямой билирубин частично попадает в большой круг кровообращения, что ведёт к желтухе. Экскреция жёлчи также нарушена. Билирубина в кишечник попадает меньше, чем в норме.
При печёночно-клеточной желтухе повышается концентрация в крови как общего билирубина, так и обеих его фракций - неконъюги-рованного (непрямого) и конъюгированного (прямого).
Так как в кишечник поступает меньше били-рубинглюкуронида, то и количество образующегося уробилиногена также снижено. Поэтому кал гипохоличньгй, т.е. менее окрашенньгй. Моча, наоборот, имеет более интенсивную окраску за счёт присутствия там не только уробилинов, но и конъюгированного билирубина, который хорошо растворим в воде и экскретируется с мочой.
3. Механическая, или обтурационная (подпечё-ночная) желтуха
Механическая, или обтурационная (подпечё-ночная), желтуха развивается при нарушении желчеотделения в двенадцатиперстную кишку. Это может быть вызвано закупоркой жёлчных протоков, например при желчнокаменной болезни, опухолью поджелудочной железы, жёлчного пузыря, печени, двенадцатиперстной кишки, хроническим воспалением поджелудочной железы или послеоперационным сужением общего жёлчного протока (рис. 13-18).
При полной закупорке общего жёлчного протока конъюгированный билирубин в составе
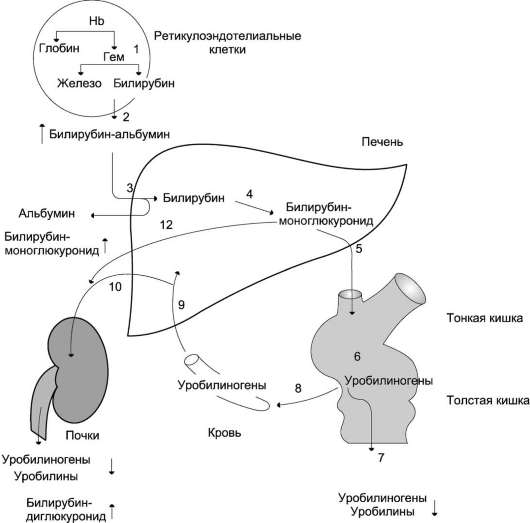
Рис. 13-17. Нарушение билирубин-уробилиногенового цикла при печёночно-клеточной желтухе. В печени снижена скорость реакции глюкуронирования билирубина (4), поэтому в крови повышается концентрация непрямого билирубина; вследствие нарушения паренхимы печени часть образованного в печени билирубинглюкуронида попадает в кровь (12) и далее с мочой (10) удаляется из организма. В моче больных присутствуют уробилины и билирубинглюкурониды. Остальные цифры соответствуют этапам метаболизма билирубина на рис. 13-16.
жёлчи не поступает в кишечник, хотя гепато-циты продолжают его вырабатывать. Поскольку билирубин в кишечник не попадает, продуктов его катаболизма уробилиногенов в моче и кале нет. Кал обесцвечен. Так как нормальные пути экскреции билирубина заблокированы, происходит его утечка в кровь, поэтому в крови больных повышена концентрация конъюгиро-ванного билирубина. Растворимый билирубин экскретируется с мочой, придавая ей насыщенный оранжево-коричневый цвет.
Б. ДИФФЕРЕНЦИАЛЬНАЯ ДИАГНОСТИКА ЖЕЛТУХ
При диагностике желтух надо иметь в виду, что на практике редко отмечают желтуху какого-либо одного типа в «чистом» виде. Чаще встречается сочетание того или иного типа. Так, при выраженной гемолитической желтухе, сопровождающейся повышением концентрации непрямого билирубина, неизбежно страдают различные органы, в том числе и печень, что может вносить элементы

Рис. 13-18. Нарушение билирубин-уробилиногенового цикла при обтурационной желтухе. Вследствие закупорки жёлчного пузыря билирубинглюкуронид не секретируется в жёлчь (5); отсутствие билирубина в кишечнике приводит к обесцвечиванию кала (6); растворимый билирубинглюкуронид выделяется почками с мочой (10). Уробилинов в моче нет; образующийся в печени билирубинглюкуронид поступает в кровь (12), вследствие этого возрастает содержание прямого билирубина. Остальные цифры соответствуют этапам метаболизма билирубина на рис. 13-16.
паренхиматозной желтухи, т.е. повышение в крови и моче прямого билирубина. В свою очередь, паренхиматозная желтуха, как правило, включает в себя элементы механической. При подпечё-ночной (механической) желтухе, например, при раке головки поджелудочной железы, неизбежен повышенный гемолиз как результат раковой интоксикации и, как следствие, повышение в крови прямого и непрямого билирубина.
Итак, гипербилирубинемия может быть следствием избытка как связанного, так и свободного
билирубина. Измерение их концентраций по отдельности необходимо при постановке диагноза желтухи. Если концентрация билирубина в плазме <100 мкмоль/л и другие тесты функции печени дают нормальные результаты, возможно предположить, что повышение обусловлено за счёт непрямого билирубина. Чтобы подтвердить это, можно сделать анализ мочи, поскольку при повышении концентрации непрямого билирубина в плазме прямой билирубин в моче отсутствует.
При дифференциальной диагностике желтух необходимо учитывать содержание уробилиноге-нов в моче. В норме за сутки из организма выделяется в составе мочи около 4 мг уробилиногенов. Если с мочой выделяется повышенное количество уробилиногенов, то это - свидетельство недостаточности функции печени, например при печёночной или гемолитической желтухе. Присутствие в моче не только уробилиногенов, но и прямого билирубина указывает на поражение печени и нарушение поступления жёлчи в кишечник.
В. НАСЛЕДСТВЕННЫЕ НАРУШЕНИЯ МЕТАБОЛИЗМА БИЛИРУБИНА
Известно несколько заболеваний, при которых желтуха вызвана наследственными нарушениями метаболизма билирубина.
Примерно у 5% населения диагностируют наследственную желтуху, вызванную генетическими нарушениями в структуре белков и ферментов, ответственных за транспорт (захват) непрямого билирубина в печень и его конъюгацию с глюку-роновой кислотой. Эта патология наследуется по аутосомно-доминантному типу. В крови больных повышена концентрация непрямого билирубина.
Известно 2 типа наследственных желтух, обусловленных нарушением реакции глюкуро-нирования в печени - образования прямого билирубина.
Для первого типа характерно полное отсутствие УДФ-глюкуронилтрансферазы. Заболева-
ние наследуется по аутосомно-рецессивному типу. Введение фенобарбитала, индуктора УДФ-глюкуронилтрансферазы, не приводит к снижению уровня билирубина. Дети умирают в раннем возрасте из-за развития билирубиновой энцефалопатии.
Для второго типа характерно снижение активности (недостаточности) УДФ-глюкуронил-трансферазы, гипербилирубинемия возникает за счёт непрямого билирубина. Желтуха хорошо поддаётся лечению фенобарбиталом.
Нарушение активного транспорта образованных в клетках печени билирубинглюкуронидов в жёлчь характерно для желтухи, наследуемой по аутосомно-доминантному типу. Проявляется гипербилирубинемией за счёт прямого билирубина и билирубинурией (в моче определяется прямой билирубин).
Семейная гипербилирубинемия новорождённых связана с наличием конкурентных ингибиторов конъюгации билирубина (эстрогенов, свободных жирных кислот) в материнском молоке. При грудном вскармливании ингибиторы конъюгации билирубина обнаруживают в сыворотке крови ребёнка. Такая гиперби-лирубинемия была названа транзиторной. Ги-пербилирубинемия исчезает при переводе ребёнка на искусственное вскармливание. Не поддающаяся лечению гипербилирубинемия приводит к развитию билирубиновой энцефалопатии и ранней смерти.
Виды порфирий - Лекция, раздел Химия, КУРС ЛЕКЦИЙ ПО ОБЩЕЙ БИОХИМИИ Острая Перемежающая Порфирия (Опп) – Причина – Дефект Ге.
Острая перемежающая порфирия (ОПП) – причина – дефект гена, кодирующего ПБГ – дезаминазу.
Наследуется по аутосомно-доминатному типу. Происходит накопление ранних предшественников синтеза гема: 5- АЛК (5- ALA) и порфобилиногена (ПБГ).
Врожденная эритропоэтическаяпорфирия - редкое врожденное заболевание, наследуемое по аутосомно-рецессивному типу. Для нее характерна низкая активность уропорфириноген-III-косинтазы и высокая - уропорфириноген-I-синтазы. Образование уропорфириногена-I значительно превосходит синтез уропорфириногена-III (нормального изомера на пути синтеза гемма). Хотя генетическое нарушение распространяется на все клетки, проявляется оно по неизвестной причине преимущественно в эритропоэтической ткани. Пациенты экскретируют с мочой большие количествауропорфириногена-I и копропорфириногена-I; которые на воздухе самопроизвольно окисляются в уропорфирин-I и копропорфирин-I - красные флуоресцирующие пигменты. Циркулирующие эритроциты содержат большое количество уропорфирина-I, однако, наивысшая концентрация этого порфирина отмечена в клетках костного мозга.
Отмечаетсясветочувствительность кожи, обусловленная активацией ПОЛ при действии света на порфириновые соединения. У пациентов отмечаются трещины на коже, часто наблюдаются гемолитические явления.
Наследственная копропорфирия - аутосомно-доминантное нарушение, обусловленное дефицитом копропорфириногеноксидазы - митохондриального фермента, ответственного за превращение копропорфириногена III в протопорфириноген IX. Копропорфириноген III в больших количествах удаляется из организма в составе фекалий и с мочой. Копропорфириноген на свету и воздухе быстро окисляется, превращаясь в красный пигмент копропорфирин.
Ограниченная способность к синтезу гема (особенно в стрессовых условиях) приводит к дерепрессии АЛК-синтазы. В результате наблюдается избыточное образование АЛК, порфобилиногена и других интермедиатов синтеза гема. У пациентов обнаруживаются все признаки и симптомы, связанные с избытком АЛК и порфобилиногена, которые характерны для перемежающейся острой порфирии, имеется повышенная светочувствительность, обусловленная присутствием избыточных количеств копропорфириногенов и уропорфириногенов.
Мозаичная порфирия, или наследственная фоторопорфирия, является аутосомно-доминантным нарушением, при котором происходит частичное блокирование ферментативного превращения протопорфириногена в гем. В норме это превращение осуществляется двумя ферментами, протопорфириногеноксидазой и феррохелатазой, локализованнымив митохондриях. Содержание протопорфириногеноксидазы составляет лишь половину нормального количества. У пациентов с мозаичной порфирией наблюдается относительная недостаточность содержания гема в стрессовых условиях, а также дерепрессированное состояние печеночной АЛК-синтазы. Что ведет к перепроизводству всех интермедиатов синтеза гема на участках перед заблокированной стадией. Пациенты с мозаичной порфирией экскретируют с мочой избыточные количества АЛК, порфобилиногена, уропорфирина и копропорфирина, а с фекалиями выделяют уропорфирин, копропорфирин и протопорфирин. Моча больных пигментирована и флуоресцирует, а кожа чувствительна к свету так же, как и у больных поздней кожной порфирией (см. ниже).
Поздняя кожная порфирия является наиболее распространенной формой порфирии. Обычно она связана с поражениями печени, особенно при избыточном потреблении алкоголя или перегрузке ионами железа. Вероятной причиной является частичная недостаточность уропорфириноген-декарбоксилазы. Нарушение, по-видимому, передается как аутосомно-доминантный признак, но генетическая пенетрантность различна и в большинстве случаев зависит от наличия нарушений функций печени. Моча содержит повышенные количества уропорфиринов типа I и III; в то же время экскреция с мочой АЛК и порфобилиногена наблюдается сравнительно редко. Иногда моча содержит весьма значительное количество порфиринов, придающих ей розоватый оттенок; при подкислении она чаще всего дает в ультрафиолетовой области розовую флуоресценцию.
Эта тема принадлежит разделу:
КУРС ЛЕКЦИЙ ПО ОБЩЕЙ БИОХИМИИ
Кафедра биохимии. КУРС ЛЕКЦИЙ ПО ОБЩЕЙ БИОХИМИИ.
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Виды порфирий
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
Все темы данного раздела:
Кровь - это жидкая ткань, которая выполняет в организме ряд важных функций: Основная функция крови - это транспорт веществ и тепловой энергии; а) Дыхательная функция.
Все жидкости организма обладают рядом общих свойств: объемом, плотностью, вязкостью, рН и осмотическим давлением. Нормальные значения общих свойств крови взрослого человека:
Форменные элементы (клетки) крови составляют 45% от общего объема крови. № Клетки крови Концентрация % от общего объема к
Химический состав растворимых в плазме крови веществ относительно постоянен, так как существуют мощные нервные и гуморальные механизмы, поддерживающие гомеостаз.
Показатель Возраст 1 день 1 мес. 6 мес. 1 год 1-6 л 12 л
В плазме крови открыто более 200 видов белков, которые составляют 7% объема плазмы. Белки плазмы крови синтезируются в основном в печени и макрофагах, а также в эндотелии сосудов, в кишечнике, лимф
По строению белки плазмы крови являются глобулярными, по составу они делятся на простые (альбумины) и сложные. Среди сложных, можно выделить липопротеины (ЛПОНП, ЛППП, ЛПНП, ЛПВП, ХМ), гли
Основным белком этой фракции является альбумин. Альбумин. Простой белок из 585 АК с массой 69кДа, имеет 17 дисульфидных мостиков, много дикарбоновых АК, обладает высок
ЛПОНП - образуются в печени. Транспорт ТГ, ХС. ЛППП - образуются в крови из ЛПОНП. Транспорт ТГ, ХС.
Синтезируются функционально активными В-лимфоцитами (плазмоцитами). У взрослого человека 107 клонов В-лимфоцитов которые синтезируют 107 видов γ-глобулинов. γ-Глобул
Понятие "белки острой фазы" объединяет до 30 белков плазмы крови, участвующих в реакции воспалительного ответа организма на повреждение. Белки острой фазы синтезируются в печени, их конце
Ферменты, находящиеся в плазме крови, можно разделить на 3 основные группы: 1. Секреторные. Они синтезируются в печени, эндотелии кишечника, сосудов поступают в кро
Факультеты: лечебно-профилактический, медико-профилактический, педиатрический. 2 курс. Эритроциты (erythrosytus) это форменные элементы крови,
Эритроциты у человека и млекопитающих в токе крови обычно (80%) имеют форму двояковогнутых дисков и называются дискоцитами. Такая форма эритроцитов создаёт наибольшую площад
Олигосахариды (сиаловая кислота и антигенные олигосахариды) гликолипидов и гликопротеидов, расположенные на наружной поверхности плазмолеммы, образуют гликокаликс.
В зрелом эритроците белки не синтезируются, т.к. у него нет рибосом, ЭПР, аппарата Гольджи и ядра. Однако в цитоплазме синтезируется пептид глутатион. Биосинтез глутатиона осуществляется в
В зрелом эритроците: 1. из ФРПФ (из рибозо-5ф) и аденина может синтезироваться АМФ. 2. АМФ с участием АТФ превращается в АДФ. 3. В реакциях субстратного фосфорилирования
В течение суток до 3% гемоглобина может спонтанно окисляться в метгемоглобин: Hb (Fe2+) ® Met Hb (Fe3+) +e- Восстановление метгемоглобина до гемог
Гем - это порфирин, в центре которого находит
Гем является простатической группой многих белков: гемоглобина, миоглобина, цитохромов митохондриальной ЦПЭ, цитохрома Р450, ферментов каталазы, пероксидазы, цитохромоксидазы, триптофанп
Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени. В костном мозге гем необходим для синтеза гемоглобина, в гепатоцитах - для образования цитохрома Р450
Синтезированный в митохондриях гем индуцируется синтез цепей глобина на полирибосомах. Гены цепей глобина расположены в 11 и 16 хромосоме. Цепи глобина формируют глобулы и соединяются с ге
Гемоглобин - тетрамерный хромопротеин, имеет массу 64,5кДа, состоит из 4 гемов и 4 глобинов. Глобины представлены полипептидными цепями различных типов a, b, g, d и т.д. a-цепь
Гемоглобин со свободной шестой координационной связью железа в составе гема называется апогемоглобином. Шестая координационная связь может связывать различные лиган
Гемоглобин присоединяет О2 последовательно, по одной молекуле на каждый гем. В апогемоглобине, благодаря координационной связи с белковой частью, атом же
Кооперативность в работе протомеров гемоглобина формирует сигмовидный характер кривой насыщения его кислородом в зависимости от парциального давления кислорода. S–образная кривая насыщения
Влияние рН на характер кривой диссоциации оксигемоглобина называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). &nb
2,3-ДФГ снижает сродство гемоглобина к кислороду и, таким образом, повышает отдачу кислорода тканям. Если кровь израсходовала весь свой запас ДФГ, гемоглобин остается фактически насыщенным кислород
1) эмбриональные гемоглобины (Gover I, Gover II). На ранних этапах развития плода в первые недели развития, когда в желточном мешке возникают очаги кроветворения начинается
Аномальные гемоглобины возникают в результате мутации генов, кодирующих a и b цепи. Известно несколько сотен мутантных гемоглобинов человека (в большинстве случаев функционально активных).
Болезни гемоглобинов называют гемоглобинозами, их насчитывают более 200. Гемоглобинозы делятся на гемоглобинопатии и таласемии. Гемоглобинопатии
Старые поврежденные эритроциты фагоцитируются клетками РЭС и перевариваются в лизосомах. При распаде гемоглобина образуется жёлчный пигмент билирубин. Дальнейший катаболизм билирубина в печени, киш
Хотите получать на электронную почту самые свежие новости?
Подпишитесь на Нашу рассылку
Новости и инфо для студентов
Реклама
Соответствующий теме материал
- Похожее
- Популярное
- Облако тегов
- Здесь
- Временно
- Пусто
О Сайте
Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
Порфирии биохимия
Гем, железосодержащее тетрагидропиррольное красящее вещество, является составной частью О 2 -связывающих белков (см. с. 274) и различных коферментов оксидоредуктаз (см. сс. 108, 310). Почти на 85% биосинтез гема происходит в костном мозге и лишь небольшая часть - в печени. В синтезе гема участвуют митохондрии и цитоплазма.
Синтез тетрагидропиррольных колец начинается в митохондриях. Из сукцинил-КоА (на схеме наверху), промежуточного продукта цитратного цикла, конденсацией с глицином получается продукт, декарбоксилирование которого приводит к 5-аминолевулинату (ALA). Отвечающая за эту стадию 5-аминолевулинат-синтаза (ALA-синтаза) [ 1 ] является ключевым ферментом всего пути. Экспрессия синтеза ALA-синтазы тормозится гемом, т. е. конечным продуктом, и имеющимся ферментом. Это типичный случай торможения конечным продуктом, или ингибирования по типу обратной связи.
После синтеза 5-аминолевулинат переходит из митохондрий в цитоплазму, где две молекулы конденсируются в порфобилиноген, который уже содержит пиррольное кольцо [ 2 ]. Порфобилиноген-синтаза [ 2 ] ингибируется ионами свинца. Поэтому при острых отравлениях свинцом в крови и моче обнаруживают повышенные концентрации 5-аминолевулината.
На последующих стадиях образуется характерная для порфирина тетрапиррольная структура. Связывание четырех молекул порфобилиногена с отщеплением NH 2 -групп и образованием уропорфириногена III катализируется гидроксиметилбилан-синтазой [ 3 ]. Для образования этого промежуточного продукта необходим второй фермент, уропорфириноген III-синтаза [ 4 ]. Отсутствие этого фермента приводит к образованию «неправильного» изомера - уропорфириногена I.
Тетрапиррольная структура уропорфиринoгена III все еще существенно отличается от гема. Так, отсутствует центральный атом железа, а кольцо содержит только 8 вместо 11 двойных связей. Кроме того, кольца несут только заряженные боковые цепи R (4 ацетатных и 4 пропионатных остатков). Так как группы гема в белках функционируют в неполярном окружении, необходимо, чтобы полярные боковые цепи превратились в менее полярные. Вначале четыре ацетатных остатка (R 1) декарбоксилируются с образованием метильных групп (5). Образующийся копропорфириноген III снова возвращается в митохондрии. Дальнейшие стадии катализируются ферментами, которые локализованы на/или внутри митохондриальной мембраны. Прежде всего под действием оксидазы две пропионатные группы (R 2) превращаются в винильные (6). Модификация боковых цепей заканчивается образованием протопорфириногена IX.
На следующей стадии за счет окисления в молекуле создается сопряженная π-электронная система, которая придает гему характерную красную окраску. При этом расходуется 6 восстановительных эквивалентов (7). В заключение с помощью специального фермента, феррохелатазы, в молекулу включается атом двухвалентного железа (8). Образованный таким образом гем или Fe-протопорфирин IX включается, например, в гемоглобин и миоглобин (см. с. 274), где он связан нековалентно, или в цитохром С, с которым связывается ковалентно (см. с. 108).
Известен ряд заболеваний, вызванных наследственными или приобретенными нарушениями порфиринового синтеза, так называемые порфирии; некоторые из них протекают очень тяжело. Многие из этих заболеваний приводят к выделению предшественников гема с калом или мочой, которая вследствие этого может быть окрашена в темно-красный цвет. Также наблюдается отложение порфиринов в коже. При воздействии света это приводит к образованию трудноизлечимых волдырей. При порфириях часты также неврологические нарушения. Возможно, что в основе средневековых легенд о людях-вампирах (дракулах) лежит странное поведение больных порфириями (светобоязнь, необычные внешность и поведение, употребление крови в пищу, компенсирующее дефицит гема и зачастую улучшающее состояние при некоторых формах порфирий).
У гемоглобина есть молекулярные болезни
Серповидно-клеточная анемия
HbS – гемоглобин серповидно-клеточной анемии. При этом нарушении в ДНК в результате точковой мутации триплет ЦТТ заменен на триплет ЦАТ, что влечет за собой включение в 6-м положении β-цепи вместо глутамата аминокислоты валина. Изменение свойств β-цепи влечет изменение свойств всей молекулы и формирование на поверхности гемоглобина «липкого» участка. При дезоксигенации гемоглобина участок «раскрывается» и связывает одну молекулу дезоксигемоглобина S с другими подобными. Результатом является полимеризация гемоглобиновых молекул и образование крупных белковых тяжей, вызывающих деформацию эритроцита и при прохождении капилляров гемолиз.
Схема отличия гемоглобина S от гемоглобина А и его полимеризация
Нарушение синтеза гемоглобина
Порфирии
Порфирии – это группа гетерогенных наследственных заболеваний, возникающих в результате нарушения синтеза гема и повышения содержания порфиринов и их предшественников в организме. Выделяют наследственные и приобретенные формы порфирии.
Приобретенные формы порфирий носят токсический характер и вызываются действием гексахлорбензола, солей свинца и других тяжелых металлов (ингибирование порфобилиногенсинтазы, феррохелатазы и др.), лекарственными препаратами (антигрибковый антибиотик гризеофульфин).
При наследственных формах дефект фермента имеется во всех клетках организма, но проявляется только в одном типе клеток. Можно выделить две большие группы порфирий:
1. Печеночные – группа заболеваний с аутосомно-доминантными нарушениями ферментов различных этапов синтеза протопорфирина IX. Наиболее ярким заболеванием этой группы является перемежающаяся острая порфирия, при которой у гетерозигот активность уропорфириноген-I-синтазы снижена на 50%.
Заболевание проявляется после достижения половой зрелости из-за повышенной потребности гепатоцитов в цитохроме Р 450 для обезвреживания половых стероидов, обострение состояния также часто бывает после приема лекарственных препаратов, метаболизм которых требует участия цитохрома Р 450 . Снижение концентрации гема, используемого для синтеза цитохрома Р 450 , активирует аминолевулинатсинтазу. В результате больные экскретируют с мочой большие количества порфобилиногена и аминолевулиновой кислоты. На свету порфириноген окисляется в окрашенные порфобилин и порфирин, и это является причиной потемнения мочи при ее стоянии на свету при доступе воздуха. Симптомами являются острые боли в животе, запоры, сердечно-сосудистые нарушения, нервно-психические расстройства.
2. Эритропоэтические – аутосомно-рецессивные нарушения некоторых ферментов синтеза протопорфирина IX в эритроидных клетках. При этом смещается баланс реакций образования уропорфириногенов в сторону синтеза уропорфириногена I. Симптомы заболевания схожи с предыдущим, но дополнительно наблюдается светочувствительность кожи, обусловленная наличием уропорфириногенов, кроме этого отмечаются трещины на коже и гемолитические явления.
Талассемии
Для талассемий характерно снижение синтеза α-цепей гемоглобина (α-талассемия) или β-цепей (β-талассемия). Это приводит к нарушению эритропоэза, гемолизу и тяжелым анемиям.
Вы можете спросить или оставить свое мнение.
Виды порфирий
Нарушения синтеза гема. Порфирии
Порфирии - гетерогенная группа заболеваний, вызванная нарушениями синтеза гема вследствие дефицита одного или нескольких ферментов.
Единой классификации порфирий нет. Порфирии делят по причинам на:
1) Наследственные . Возникают при дефекте гена фермента, участвующего в синтезе гема;
2) Приобретенные . Возникают при ингибирующем влиянии токсических соединений (гексохлорбензол, соли тяжелых металлов - свинец) на ферменты синтеза гема.
В зависимости от преимущественной локализации дефицита фермента (в печени или эритроцитах) порфирин делится на:
1) печеночные – наиболее распространенный тип порфирина к нему относится острая перемежающаяся порфирия (ОПП), поздняя кожная порфирия, наследственная копропорфирия, мозаичная порфирия;
2) эритропоэтические – врожденная эритропоэтическая порфирия (болезнь Гюнтера), эритропоэтическая протопорфирия.
В зависимости от клинической картины, порфирии делят на:
Негативные последствия порфирий связаны с дефицитом гема и накоплением в тканях и крови промежуточных продуктов синтеза гема – порфириногенов и продуктов их окисления. При эритропоэтических порфириях порфирины накапливаются в нормобластах и эритроцитах, при печёночных - в гепатоцитах.
Для каждого вида порфирии существует определенный уровень ферментативного дефекта, в результате накапливаются продукты, синтезирующиеся выше этого уровня. Эти продукты являются основными диагностическими маркерами заболевания.
Порфириногены ядовиты, при тяжёлых формах порфирий они вызывают нейропсихические расстройства, нарушения функций РЭС и повреждения кожи.
Нейропсихические расстройства при порфириях связаны с тем, что аминолевулинат и порфириногены являются нейротоксинами.
В коже на солнце порфириногены легко превращаются в порфирины. Кислород при взаимодействии с порфиринами переходит в синглетное состояние. Синглетный кислород стимулирует ПОЛ клеточных мембран и разрушение клеток, поэтому порфирии часто сопровождаются фотосенсибилизацией и изъязвлением открытых участков кожи.
Порфириногены не окрашены и не флуоресцируют, а порфирины проявляют интенсивную красную флуоресценцию в ультрафиолетовых лучах. Избыток порфиринов который выводиться с мочой, придает ей темный цвет («порфирин» в переводе с греч. означает пурпурный).
Иногда при лёгких формах наследственных порфирии заболевание может протекать бессимптомно, но приём лекарств, являющихся индукторами синтеза аминолевулинатсинтазы, может вызвать обострение болезни. В некоторых случаях симптомы болезни не проявляются до периода полового созревания, когда повышение образования β-стероидов вызывает индукцию синтеза аминолевулинатсинтазы. Порфирии наблюдают и при отравлениях солями свинца, так как свинец ингибирует аминолевулинатдегидратазу и феррохелатазу. Некоторые галогенсодержащие гербициды и инсектициды являются индукторами синтеза аминолевулинатсинтазы, поэтому попадание их в организм сопровождается симптомами порфирии.
Острая перемежающая порфирия (ОПП) – причина – дефект гена, кодирующего ПБГ – дезаминазу. Наследуется по аутосомно-доминатному типу. Происходит накопление ранних предшественников синтеза гема: 5- АЛК (5- ALA) и порфобилиногена (ПБГ).
Бесцветный ПБГ на свету превращается в порфибилин и порфирин, они предают моче темный цвет. АЛК оказывает нейротоксическое действие, приводя к вялому параличу конечностей и парезу дыхательной мускулатуры. Последнее вызывает острую дыхательную недостаточность. Заболевание проявляется в среднем возрасте, провоцируется приемом анальгетиков, сульфаниломидных препаратов, так как они увеличивают синтез АЛК – синтазы.
Клинической симптоматикой являются острые боли в животе, рвота, запор, сердечно-сосудистые нарушения, нервно-психические расстройства. Не наблюдается повышенной чувствительности к свету, так как метаболическое нарушение проходит на стадии, предшествующей образованию уропорфириногена.
Для лечения применяют препарат нормосанг – аргинат гема. Действие основано на том, что гем, по механизму отрицательной обратной связи блокирует трансляцию АЛК – синтазы, а, следовательно, падает синтез АЛК и ПБГ, чем и достигается купирование симптоматики.
Врожденная эритропоэтическаяпорфирия - это еще более редкое врожденное заболевание, наследуемое по аутосомно-рецессивному типу. Молекулярная природа этой болезни точно неизвестна; установлено, однако, что для нее характерен определенный дисбаланс относительных активностей уропорфириноген-Ш-косинтазы и уропорфириноген-1-синтазы. Образование уропорфириногена I в количественном отношении значительно превосходит синтез уропорфириногена III-нормального изомера на пути синтеза гема. Хотя генетическое нарушение распространяется на все клетки, проявляется оно по неизвестной причине преимущественно в эритропоэтической ткани. Пациенты с врожденной эритропоэтической порфирией экскретируют большие количестваизомеров типа I уропорфириногена и копропорфириногена; в моче оба этих соединения самопроизвольно окисляются в уропорфирин I и копропорфирин I-красные флуоресцирующие пигменты. Сообщалось о случае, когда наблюдалось небольшое повышение концентрации уропорфирина III, но отношение изомеров типа I и III составляло примерно 100:1. Циркулирующие эритроциты содержат большое количество уропорфирина 1, однако, наивысшая концентрация этого порфирина отмечена в клетках костного мозга (но не в гепатоцитах).
Отмечаетсясветочувствительность кожи, обусловленная характером спектра поглощения порфириновых соединений, которые образуются в больших количествах. У пациентов отмечаются трещины на коже, часто наблюдаются гемолитические явления.
Наследственная копропорфирия - аутосомно-доминантное нарушение, обусловленное дефицитом копропорфнрнногеноксидазы - митохондриального фермента, ответственного за превращение копропорфириногена III в протопорфириноген IX. Копропорфириноген III в больших количествах удаляется из организма в составе фекалий, а также вследствие его растворимости в воде экскретируется в большом количестве с мочой. Как и уропорфириноген, копропорфириноген на свету и воздухе быстро окисляется, превращаясь в красный пигмент копропорфирин.
Ограниченная при этом заболевании способность к синтезу гема (особенно в стрессовых условиях) приводит к дерепрессии АЛК-сиитазы. В результате наблюдается избыточное образование АЛК и порфобилиногена, а также других интермедиатов на пути синтеза тема, образующихся на стадиях, предшествующих наследственно заблокированному этапу. Соответственно у пациентов с наследственной копропорфирией обнаруживаются все признаки и симптомы, связанные с избытком АЛК и порфобилиногена, которые характерны для перемежающейся острой порфирии, но помимо этого у них имеется повышенная светочувствительность, обусловленная присутствием избыточных количеств копропорфириногенов и уропорфириногенов. При этом заболевании введение гематина также может вызвать по крайней мере частичную репрессию АЛК-синтазы и смягчение симптомов, обусловленных перепроизводством интермедиатов биосинтеза гема.
Мозаичная порфирия, или наследственная фоторопорфирия, является аутосомно-доминантным нарушением, при котором происходит частичное блокирование ферментативного превращения протопорфириногена в гем. В норме это превращение осуществляется двумя ферментами, протопорфириногеноксидазой и феррохелатазой, локализованнымив митохондриях. Судя по данным, полученным на культуре фибробластов кожи, у больных мозаичной порфирией содержание протопорфириногеноксидазы составляет лишь половину нормального количества. У пациентов с мозаичной порфирией наблюдается относительная недостаточность содержания гема в стрессовых условиях, а также дерепрессированное состояние печеночной АЛК-синтазы. Как отмечалось выше, повышенная активность АЛК-синтазы ведет к перепроизводству всех интермедиатов синтеза гема на участках перед заблокированной стадией. Таким образом, пациенты с мозаичной порфирией экскретируют с мочой избыточные количества АЛК, порфобилиногена, уропорфирина и копропорфирина, а с фекалиями выделяют уропорфирин, копропорфирин и протопорфирин. Моча больных пигментирована и флуоресцирует, а кожа чувствительна к свету так же, как и у больных поздней кожной порфирией (см. ниже).
Поздняя кожная порфирия, вероятно, является наиболее распространенной формой порфирии. Обычно она связана с теми или иными поражениями печени, особенно при избыточном потреблении алкоголя или перегрузке ионами железа. Природа метаболического нарушения точно не установлена, но вероятной причиной является частичная недостаточность уропорфириноген-декарбоксилазы. Нарушение, по-видимому, передается как аутосомно-доминантный признак, но генетическая пенетрантность различна и в большинстве случаев зависит от наличия нарушений функций печени. В соответствии с предсказаниями моча содержит повышенные количества уропорфиринов типа I и III; в то же время экскреция с мочой АЛК и порфобилиногена наблюдается сравнительно редко. Иногда моча содержит весьма значительное количество порфиринов, придающих ей розоватый оттенок; при подкислении она чаще всего дает в ультрафиолетовой области розовую флуоресценцию.
Печень содержит большие количества порфиринов и поэтому сильно флуоресцирует, тогда как у эритроцитов и клеток костного мозга флуоресценция отсутствует. Главным клиническим проявлением при поздней кожной порфирии является повышеннаясветочувствительность кожи. У больных не наблюдается ни повышенной активности АЛК-синтазы, ни соответственно избыточного содержания в моче порфобилиногена и АЛК; это коррелирует с отсутствием острых приступов, характерных для перемежающейся острой порфирии.
Протопорфирия, или эритропоэтическая протопорфирия, по-видимому, обусловлена доминантно наследуемой недостаточной активностьюферрохелатазы в митохондриях всех тканей; клинически эта болезнь проявляется как острая крапивница, вызываемая воздействием солнечных лучей. Эритроциты, плазма и фекалии содержат повышенные количества протопорфирина IX, а ретикулоциты (незрелые эритроциты) и кожа (при исследовании с помощью биопсии) часто флуоресцируют красным светом. Печень, вероятно, тоже вносит вклад в повышение образования протопорфирина IX, однако экскреции с мочой порфиринов и их предшественников не наблюдается.
Порфирии биохимия
Порфириями называют гетерогенную группу заболеваний, характеризующихся повышенным выделением порфиринов или их предшественников. Некоторые формы порфирии являются наследственными, другие - приобретенными. Было предложено несколько различных классификаций порфирий. Наследственные формы удобно разделить на три большие группы - эритропоэтическне, печеночные и такие формы, при которых нарушения метаболизма наблюдаются одновременно в эритропоэтической и печеночной тканях (табл. 33.2). Для большинства наследуемых форм характерно наличие метаболических
Таблица 33.2. Классификация порфирий человека
лических нарушений во всех тканях, однако проявляются они по каким-то причинам в каком-то одном типе тканей. Ниже приводится описание биохимических нарушений, характерных для порфирий.
Для каждого типа порфирии характерен набор экскретируемых с мочой порфиринов и их предшественников. Эти данные и их связь с различными стадиями синтеза гема приведены на рис. 33.11.
Перемежающаяся острая порфнрня (ПОП) является признаком аутосомнодоминантной наследственной болезни человека, которая обычно проявляется только после достижения половой зрелости. Ее причиной является наследуемая частичная недостаточность уропорфириноген-1-синтазы. Больные гетерозиготны по дефектному структурному гену, поэтому активность уропорфириноген-1-синтазы в их клетках составляет 50% от нормы. Пациенты с ПОП экскретируют с мочой большие количества порфобилиногена и АЛК. Оба этих соединения бесцветны, но порфобилиноген на свету и воздухе самопроизвольно образует два окрашенных продукта - порфобилин и порфирин. Это является причиной потемнения мочи при ее стоянии на свету при доступе воздуха.
Порфобилиноген и АЛК присутствуют в плазме и спинномозговой жидкости больных, особенно в период резкого обострения. Лекарственные препараты и стероидные гормоны, метаболизм которых требует участия гемсодержащих белков, таких, как цитохром Р-450, могут ускорять наступление обострения. Соединения, индуцирующие порфирию в процессе метаболизма, повышают потребление гемовых белков и тем самым снижают внутриклеточную концентрацию гема; это приводит к дерепрессии синтеза АЛК-синтазы. Повышение активности АЛК-синтазы и частичное блокирование уропорфириноген-1-синтазы приводит к значительному накоплению АЛК и порфобилиногена; это сопровождается острыми болями в животе, рвотой, запором, сердечно-сосудистыми нарушениями, а также нервно-психическими расстройствами. Следует отметить, что, согласно экспериментальным данным, снижение содержания гема угнетает активность триптофанпирролазы и приводит к накоплению нейроактивных соединений - триптофана и 5-гидрокситриптамина.
У пациентов с ПОП не наблюдается повышенной чувствительности к свету, которая характерна для других печеночных порфирий. Этого следовало ожидать, поскольку у больных не накапливаются ни порфирины, ни порфириногены, так как метаболическое нарушение синтеза гема происходит на стадии, предшествующей образованию первого порфириногена (уропорфириногена).
Как отмечено выше, метаболические нарушения обнаруживаются и в других клетках, в частности, в эритроцитах, а также в выращенных в культуре фибробластах или клетках амниотической жидкости; однако повышенная активность АЛК-синтазы, приводящая к перепроизводству АЛК и порфобилиногена, наблюдается преимущественно в печени. Видимо, это связано с тем, что печень является тем самым органом, в котором протекает метаболизм индуцирующих агентов. Острая порфирия служит одним из редких примеров болезни, фенотипически выраженной у гетерозигот, при этом недостаточность фермента составляет только 50%.
Как и можно было предсказать на основании
Рис. 33.11. Последовательные стадии биосинтеза гема, с указанием предшественников, экскретируемых с мочой, при различных формах порфирии Фигурные скобки объединяют соединения, которые экскретируются с мочой в избыточных количествах при обострении указанных форм порфирии. АЛК - 5-аминолевулиновая кислота. (Воспроизведено с изменениями из статьи Kaufman L., Merver Н. S. Biochemical defects in two types of human hepatic prphyria. N. Engl. J. Med 1970:283:954.)
предложенного механизма регуляции синтеза АЛК-синтазы системой репрессии - дерепрессин, введение гематина пациентам с ПОП может уменьшить индукцию АЛК-синтазы и тем самым облегчить протекание заболевания.
Врожденная эрнтропоэтическая порфирия - это еще более редкое врожденное заболевание, наследуемое по аутосомнорецессивному типу. Молекулярная природа этой болезни точно неизвестна; установлено, однако, что для нее характерен определенный дисбаланс относительных активностей уропорфириноген-III- косинтазы и уропорфириноген-1-синтазы. Образование уропорфириногена I в количественном отношении значительно превосходит синтез уропорфириногена III - нормального изомера на пути синтеза гема. Хотя генетическое нарушение распространяется на все клетки, проявляется оно по неизвестной причине преимущественно в эритропоэтической ткани. Пациенты с врожденной эритропоэтической порфирией экскретируют большие количества изомеров типа I уропорфириногена и копропорфириногена; в моче оба этих соединения самопроизвольно окисляются в уропорфирин 1 и копропорфирин I - красные флуоресцирующие пигменты. Сообщалось о случае, когда наблюдалось небольшое повышение концентрации уропорфирина III, но отношение изомеров типа I и III составляло примерно 100:1. Циркулирующие эритроциты содержат большое количество уропорфирина I, однако наивысшая концентрация этого порфирина отмечена в клетках костного мозга (но не в гепатоцитах).
Вероятно, вследствие образования меньших количеств истинного предшественника гема, уропорфириногена III, и возникающего вследствие этого относительного дефицита гема в эритропоэтических тканях больных происходит индукция АЛК-синтазы. Эта индукция приводит к перепроизводству порфириногенов типа I. Наряду с усилением синтеза АЛК-синтазы и перепроизводством порфириногенов типа I повышаются образование и экскреция порфобилиногена и АЛК. Таким образом, на основе биохимических отклонений можно предсказывать появление клинических симптомов, сходных с теми, которые наблюдаются при ПОП, но помимо этого отмечается светочувствительность кожи,
обусловленная характером спектра поглощения порфириновых соединений, которые образуются в больших количествах. У пациентов отмечаются трещины на коже, часто наблюдаются гемолитические явления.
Наследственная копропорфирия-аутосомно - доминантное нарушение, обусловленное дефицитом копропорфнрнноген-оксндазы - митохондриального фермента, ответственного за превращение копропорфириногена III в протопорфириноген IX. Копро-порфириноген III в больших количествах удаляется из организма в составе фекалий, а также вследствие его растворимости в воде экскретируется в большом количестве с мочой. Как и уропорфириноген, копропорфириноген на свету и воздухе быстро окисляется, превращаясь в красный пигмент копропорфирин.
Ограниченная при этом заболевании способность к синтезу гема (особенно в стрессовых условиях) приводит к дерепрессии АЛК-синтазы. В результате наблюдается избыточное образование АЛК и порфобилиногена, а также других интермедиатов на пути синтеза гема, образующихся на стадиях, предшествующих наследственно заблокированному этапу. Соответственно у пациентов с наследственной копропорфирией обнаруживаются все признаки и симптомы, связанные с избытком АЛК и порфобилиногена, которые характерны для перемежающейся острой порфирии, но помимо этого у них имеется повышенная светочувствительность, обусловленная присутствием избыточных количеств копропорфириногенов и уропорфириногенов. При этом заболевании введение гематина также может вызвать по крайней мере частичную репрессию АЛК-синтазы и смягчение симптомов, обусловленных перепроизводством интермедиатов биосинтеза гема.
Мозаичная порфнрня, или наследственная фотокопропорфирия, является аутосомно-доминантным нарушением, при котором происходит частичное блокирование ферментативного превращения про-топорфириногена в гем. В норме это превращение осуществляется двумя ферментами, протопорфири-ногеноксидазой и феррохелатазой, локализованными в митохондриях. Судя по данным, полученным на культуре фибробластов кожи, у больных мозаичной порфирией содержание протопорфириногеноксидазы составляет лищь половину нормального количества. У пациентов с мозаичной порфирией наблюдается относительная недостаточность содержания гема в стрессовых условиях, а также дерепрессированное состояние печеночной АЛК-синтазы. Как отмечалось выше, повышенная активность АЛК-синтазы ведет к перепроизводству всех интермедиатов синтеза гема на участках перед заблокированной стадией. Таким образом, пациенты с мозаичной порфирией экскретируют с мочой избыточные количества АЛК, порфобилиногена, уропорфирина и копропорфирина, а с фекалиями выделяют уропорфирин, копропорфирин и протопорфирин. Моча больных пигментирована и флуоресцирует, а кожа чувствительна к свету так же, как и у больных поздней кожной порфирией (см. ниже).
Поздняя кожная порфнрня, вероятно, является наиболее распространенной формой порфирии. Обычно она связана с теми или иными поражениями печени, особенно при избыточном потреблении алкоголя или перегрузке ионами железа. Природа метаболического нарушения точно не установлена, но вероятной причиной является частичная недостаточность уропорфнрнноген - декарбоксилазы. Нарушение, по-видимому, передается как аутосомно-доминантный признак, но генетическая пенетрантность различна и в большинстве случаев зависит от наличия нарушений функций печени. В соответствии с предсказаниями моча содержит повышенные количества уропорфиринов типа I и III; в то же время экскреция с мочой АЛК и порфобилиногена наблюдается сравнительно редко. Иногда моча содержит весьма значительное количество порфиринов, придающих ей розоватый оттенок; при подкислении она чаще всего дает в ультрафиолетовой области розовую флуоресценцию.
Печень содержит большие количества порфиринов и поэтому сильно флуоресцирует, тогда как у эритроцитов и клеток костного мозга флуоресценция отсутствует. Главным клиническим проявлением при поздней кожной порфирии является повышенная светочувствительность кожи. У больных не наблюдается ни повышенной активности АЛК-синтазы, ни соответственно избыточного содержания в моче порфобилиногена и АЛК; это коррелирует с отсутствием острых приступов, характерных для перемежающейся острой порфирии.
Протопорфирия, или эритропоэтическая протопорфирия, по-видимому, обусловлена доминантно наследуемой недостаточной активностью феррохелатазы в митохондриях всех тканей; клинически эта болезнь проявляется как острая крапивница, вызываемая воздействием солнечных лучей. Эритроциты, плазма и фекалии содержат повышенные количества протопорфирина IX, а ретикулоциты (незрелые эритроциты) и кожа (при исследовании с помощью биопсии) часто флуоресцируют красным светом.
Печень, вероятно, тоже вносит вклад в повышение образования протопорфирина IX, однако экскреции с мочой порфиринов и их предшественников не наблюдается.
Приобретенная (токсическая) порфнрня может быть вызвана действием токсических соединений, таких, как гексахлорбензол, соли свинца и других тяжелых металлов, а также лекарственными препаратами, например гризеофульвином. Тяжелые металлы являются ингибиторами ряда ферментов в системе синтеза гема, включая АЛК-дегидратазу, уропорфириноген - синтазу и феррохелатазу.
Раздел I. Структура и функции белков и ферментов
Раздел II. Биоэнергетика и метаболизм углеводов и липидов
Раздел III. Метаболизм белков и аминокислот
Копирование информации со страницы разрешается только с указанием ссылки на данный сайт
Урс А. Мейер (Urs А. Меуег)
Порфирии представляют собой патологию, связанную с наследственными или приобретенными аномалиями биосинтеза гема. Порфирины, эти тетрапирроловые пигменты, выполняют роль промежуточных продуктов этого пути и образуются из предшественников - 8 -аминолевулиновой кислоты (АЛК) и порфобилиногена. Гем, комплекс двухвалентного железа с протопорфирином IX, функционирует в качестве простетической группы гемопротеинов, таких как гемоглобин, цитохромы, каталаза и триптофаноксигеназа. Его биосинтез жизненно важен и происходит во всех аэробных клетках.
Каждая порфирия характеризуется особенностями гиперпродукции, накопления и экскреции промежуточных продуктов биосинтеза гема. Эти особенности отражают метаболическую экспрессию дефицита отдельных ферментов биосинтеза.
Основные клинические проявления заключаются в перемежающихся приступах дисфункции нервной системы и/или чувствительности кожи к солнечным лучам. Неврологический синдром обычно провоцируется приемом таких препаратов, как барбитураты, и заключается в болях в животе, периферической нейропатии и нарушениях психики. Нейропсихические симптомы появляются только при порфириях, при которых резко усилена продукция предшественников порфиринов - АЛК и порфобилиногена. Патогенез неврологических нарушений неясен. Светочувствительность кожи связана непосредственно с повышенным накоплением порфиринов, хотя при разных нарушениях кожные проявления неодинаковы. Светочувствительность обусловлена фотодинамическим действием порфиринов и опосредуется, вероятно, образующимся синглет-кислородом с последующим развитием деструктивных процессов, таких как перекисное окисление липидов лизосомных мембран. Доминантно наследуемые порфирии человека экспрессируются по-разному. Могут определяться лишь биохимические или ферментативные изменения. Подобное латентное течение болезни может оказаться одной из стадий или продолжается в течение всей жизни больного. В других случаях симптоматика может провоцироваться лекарственными препаратами, гормонами или повреждением печени.
Классификация. Порфирии подразделяют обычно на две основные группы (эритропоэтическая и печеночная) в соответствии с основными местами синтеза гема, в которых проявляются «ошибки» метаболизма. Единственная чисто эритропоэтическая форма порфирии - врожденная эритропоэтическая (ВЭП) - встречается редко. При протопорфирии (ПрП) порфирины накапливаются как в клетках эритроидного ряда, так и в печеночной ткани. При интермиттирующей острой порфирии, наследственной копропорфирии и пестрой порфирии (соответственно ИОП, НКП и ПП) доминантно наследуемая недостаточность ферментов обусловливает нарушение биосинтеза гема преимущественно в печени без видимых нарушений образования гемоглобина. Хроническую кожную порфирию (ХКП) ранее считали приобретенной печеночной. Однако у большинства (если не у всех) больных обнаруживают наследственную недостаточность уропорфириногендекарбоксилазы. Приобретенную порфирию, напоминающую ХКП, обусловливают воздействие полихлорированных углеводородов и опухоли печени. Отравление свинцом также сопровождается нарушением синтеза порфиринов и гема. Некоторое усиление экскреции порфиринов или их предшественников с мочой, равно как и накопление порфиринов в эритроцитах, может сопровождать многие клинические состояния. При вторичных феноменах симптомы и признаки порфирии отсутствуют.
Биохимические аспекты. Последовательность реакций синтеза гема из субстратов глицина и сукцинил-фермента А через АЛК и порфобилиноген (ПБК) катализируется четырьмя митохондриальными и четырьмя цитозольными ферментами. В регуляции биосинтеза гема в разных тканях существуют различия.
В печени скорость образования гема ограничивается реакцией, катализируемой АЛК-синтазой. Ферменты, действующие после АЛК-синтазы, определяются в избытке. Главным регулятором АЛК-синтазы служит конечный продукт всего пути - гем, который репрессирует фермент по механизму отрицательной обратной связи. Повышенные потребности в геме удовлетворяются путем новообразования АЛК-синтазы. Ее синтез в печени индуцируется большим числом жирорастворимых веществ, стероидами и химическими соединениями, которые служат субстратами и индукторами гемопротеиновых цитохромов Р 450 - конечных оксидаз на пути микросомного метаболизма фармакологических средств. Эта индукция модулируется многочисленными генетическими, метаболическими и факторами окружающей среды. При порфириях, при которых симптомы провоцируются некоторыми лекарственными препаратами, взаимозависимость синтеза гема и микросомального окисления этих препаратов приобретает большое значение.
В клетках костного мозга, в которых происходит полный синтез гема, ограничивающая скорость реакция также катализируется АЛК-синтазой, но о ее роли в синтезе гема во время деления, дифференцировки и созревания клеток эритроидного ряда известно мало. В процессе созревания этих клеток из них исчезают ядра и митохондрии и, следовательно, митохондриальные ферменты синтеза гема, тогда как цитозольные ферменты, катализирующие реакции между АЛ К и копро-порфириногеном, сохраняются. В связи с этим эритроциты можно использовать для диагностики порфирий, связанных с дефектом только цитозольного фермента.
Регуляция синтеза гема в костном мозге и печени различна. В печени основной детерминантой образования гема служит уровень АЛК-синтазы, тогда как в костном мозге синтез гема запускается сложным процессом дифференцировки эритроидной клетки. Именно поэтому, вероятно, дефекты ферментов синтеза гема в эритроидных клетках и печени проявляются по-разному.
Порфириногены занимают промежуточное положение между порфобилиногеном и протопорфирином. Они бесцветны и не флюоресцируют. За исключением протопорфирина, порфирины - это побочные продукты, которые покидают путь биосинтеза вследствие необратимого окисления соответствующего порфириногена. Порфирины не выполняют физиологической функции, но в силу своей окраски и флюоресценции определяют необычный цвет мочи и эритроцитов у некоторых больных.
От расположения двух замещенных боковых цепей на пирроловом кольце порфиринов зависят структурные типы изомеров, которые нумеруются от I до IV. В природе найдены только типы I и III, причем только тип III служит субстратом конечных этапов реакции, ведущей к образованию протопорфирина IX и гема. При распаде гема образуются не порфирины, а нециклические тетрапирролы, называемые желчными пигментами.
Врожденная эритропоэтическая порфирия
Определение. Врожденная эритропоэтическая порфирия (ВЭП, болезнь Гюнтера, врожденная светочувствительная порфирия, эритропоэтическая уропорфирия) - это редкое, рецессивно наследуемое заболевание, проявляющееся хронической светочувствительностью с обезображивающими повреждениями кожи и гемолитической анемией.
Генетика, частота и патогенез. Больные представляют собой гомозигот по аутосомно-рецессивному гену. У гетерозигот порфириновый обмен нарушается редко, внешне они выглядят здоровыми. Лежащий в основе болезни ферментный дефект не установлен, но может заключаться в функциональном неравновесии активностей порфобилиногендезаминазы и косинтазы уропорфириногена III . Эта аномалия экспрессируется исключительно в созревающих клетках эритроидного ряда и приводит к резкой гиперпродукции уропорфириногена I , тогда как продукция уропорфириногена III не изменяется или несколько повышена. Уропорфириноген I не может использоваться для синтеза гема, но превращается в копропорфириноген I . Уропорфирин I , копропорфириноген I и копропорфирин I накапливаются в тканях и в избыточных количествах экскретируются с мочой и калом.
Клинические проявления и диагностика. У больных порфирины накапливаются еще в период внутриутробного развития. Уже в родах или вскоре после них обычно начинает отделяться розовая или красная моча, тогда как светочувствительность кожи, периодический гемолиз и спленомегалия могут появиться позднее. Часто определяют гипертрихоз и окрашивание зубов и костей в красный цвет. Смерть может наступить уже в детстве. При более длительном выживании у больного появляются большие калечащие рубцы, особенно на коже пальцев рук, носа и ушей. В моче определяют большие количества уропорфирина I, копропорфирина и порфиринов с 7, 6, 5 и 3 карбоксильными группами, тогда как экскреция АЛ К и ПБГ не изменяется. В кале обнаруживают большие количества копропорфирина I. Нормобласты, ретикулоциты и эритроциты содержат большие количества уропорфирина I и малое количество копропорфириногена I . Нормобласты и ретикулоциты проявляют интенсивно красную флюоресценцию. В соответствии с нормальной экскрецией АЛ К и ПБГ неврологическая патология отсутствует.
Лечение. Необходимо исключить воздействие солнечного света. В некоторых случаях гемолитическая анемия, экскреция порфиринов и светочувствительность уменьшаются после спленэктомии. Применение инфузий гематина и перорального введения р -каротина пока не вышло за рамки эксперимента.
Печеночные порфирии
Три печеночные порфирии (ИОП, НКП и ПП) во многом сходны. Все они наследуются как аутосомный доминантный признак. Острые приступы угрожающей жизни неврологической патологии провоцируются разнообразными препаратами, гормонами и другими факторами, во время которых с мочой экскретируются большие количества АЛК и ПБГ, но виды порфиринов в моче и кале различны.
Интермиттирующая острая порфирия. Определение. Интермиттирующая острая порфирия [ИОП, острая интермиттирующая порфирия (ОИП), пирролопорфирия] характеризуется повторными приступами неврологических и психических симптомов. Светочувствительность отсутствует. Первично нарушается функция порфобилиногендезаминазы.
Генетика, частота и патогенез. Аномалия наследуется как аутосомный доминантный признак с непостоянной экспрессивностью. Аномальный ген встречается с частотой 1:10000-1:50000, но в некоторых регионах она может быть выше. Гомозиготы не встречались. Причина болезни заключается в частичной (50 %) недостаточности порфобилиногендезаминазы, превращающей ПБГ в уро-порфириноген I. На генном уровне эта недостаточность может обусловливаться не одним, а несколькими механизмами, но наиболее частая мутация обусловливает уменьшение количества иммунореактивного фермента белка. В печени частичная недостаточность фермента приводит к повышению активности и/или индуцибельности АЛК-синтазы лекарственными средствами и другими факторами и, следовательно, к повышению образования и экскреции с мочой АЛК и ПБГ. В этих условиях порфирины не накапливаются, и поэтому светочувствительность кожи не повышена. При ИОП сниженная активность порфобилиногендезаминазы определяется в печени, эритроцитах, культуре кожных фибробластов, лейкоцитах и клетках амниотической жидкости. Таким образом, ферментный дефект имеет место и во внепеченочных тканях, но его метаболические последствия в них не проявляются. Недостаточность фермента при отсутствии приобретенных факторов необязательно приводит к клинически выраженной острой порфирии, и только у "/з больных и даже менее, страдающих этим генетическим дефектом, когда-либо возникает приступ порфирии. Связь между генетическим дефектом и неврологическими нарушениями остается невыясненной.
Клинические проявления и диагностика. Симптомы болезни редко появляются до пубертатного возраста. Обычно первым и наиболее ярким симптомом приступа порфирии служит боль в животе. Она может быть умеренной или очень сильной, коликообразной, локализованной или генерализованной, иррадиирует в спину или поясницу. Боль связана, вероятно, с автономной нейропатией, сопровождающейся нарушением двигательной активности желудочно-кишечного тракта с чередующимися спазматическими и расширившимися участками кишечника. Живот обычно мягкий, и боль при надавливании не усиливается. Из-за часто сопутствующих лихорадочного состояния и лейкоцитоза острый приступ порфирии может имитировать любой воспалительный процесс в брюшной полости. Часты выраженная рвота и запор. Неврологические и психические аномалии проявляются по-разному. Могут нарушаться функции периферических нервов, автономной нервной системы, ствола мозга, черепных нервов или головного мозга. Нередки тахикардия и лабильная гипертензия с постуральной гипотензией, задержка мочи и чрезмерная потливость. Гипертензия и тахикардия коррелируют с повышенной экскрецией катехоламинов. Периферическая нейропатия обусловливается вовлечением в процесс преимущественно двигательных нервов, но может присоединиться и чувствительный компонент. Глубокие сухожильные рефлексы снижены или отсутствуют. Типичны невритные боли в конечностях, области гипо- и парестезий, а также вялое свисание стоп и кистей. Может развиться параплегия или полная вялая квадриплегия. В прошлом основной причиной смерти служил паралич дыхательной мускулатуры. При вовлечении в процесс черепных нервов могут атрофироваться зрительный нерв, присоединяться офтальмоплегия и дисфагия. При более выраженных повреждениях ЦНС появляются бред, кома и судороги. Несмотря на обратимость нейропатии, остаточные парезы могут сохраняться в течение ряда лет после острого приступа. Многие больные в течение продолжительного времени остаются раздражительными, эмоционально неустойчивыми с сохраняющимися функциональными нарушениями. У 1/3 больных нарушается психика, могут развиться органический мозговой синдром с беспокойством, дезориентацией и зрительными галлюцинациями. Иногда определяется выраженная гипонатриемия. Это может быть обусловлено несколькими причинами (в том числе выведением натрия через желудочно-кишечный тракт, неоправданным введением жидкости и сольтеряющей формой нефропатии из-за токсического эффекта АЛК), но ведущей служит, по-видимому, неадекватная секреция антидиуретического гормона. В ряде случаев присоединяется столь выраженная гипомагниемия, что развивается тетания.
Острые приступы продолжаются в течение нескольких дней и даже месяцев и варьируют по частоте и тяжести. В периоды ремиссий симптомы болезни ослабляются или полностью исчезают. Клинические (и биохимические) проявления могут провоцироваться обычными (терапевтические) дозами барбитуратов, противосудорожных средств, эстрогенами, контрацептивами или алкоголем. Все эти вещества окисляются гемопротеинами системы цитохрома Р450. Во время острых приступов метаболизм некоторых из них в печени может нарушаться. У некоторых женщин ухудшение состояния коррелирует с менструальным циклом и латентная порфирия может стать явной в поздние сроки беременности или вскоре после родов. Приступы могут провоцироваться и длительным периодом сниженного потребления калорий (голодание).
Лабораторные данные. Для острых приступов характерна избыточная экскреция с мочой АЛК и ПБГ, причем по этому признаку ИОП не отличается от НКП или ПП. Уровень АЛК и ПБГ в моче не коррелирует с выраженностью симптоматики. Простым и надежным скрининг-тестом, помогающим диагностировать острый приступ ИОП, НКП и ПП, служит качественное определение порфо-билиногена в моче (тесты Уотсона-Шварца или Хоша). При нейропсихических нарушениях эти тесты почти всегда положительны, однако для этого требуется, чтобы концентрация ПБГ в моче превышала верхнюю границу нормы в 3-5 раз. В связи с этим оба теста при латентной форме болезни или нормализации экскреции ПБГ после купирования приступа могут быть отрицательными. Иногда требуется количественное определение экскретируемых с мочой АЛК и ПБГ с помощью хроматографических методов. При латентной форме ИОП с нормальной экскрецией АЛК и ПБГ диагноз может быть установлен на основании результатов определения активности порфобилиногендезаминазы в эритроцитах, лейкоцитах или культивируемых фибробластах кожи. Однако у здоровых и больных ИОП эти результаты перекрываются, и точно поставить диагноз не всегда возможно.
При ИОП в соответствии с ферментным дефектом усилена экскреция предшественников порфиринов - АЛК и ПБГ, поэтому свежеполученная моча обычно бесцветна, содержит мало предобразованного уро- или копропорфирина. При стоянии она может темнеть, так как ПБГ спонтанно полимеризуется в уропорфирин и порфобилин - темно-коричневый пигмент неизвестного строения. У некоторых больных, однако, определяют достаточное количество образующихся неферментативным путем пигментов, чтобы придавать свежеполученной моче темно-красный цвет. Концентрация порфиринов в кале обычно находится в пределах нормы.
Общепринятые функциональные печеночные пробы не изменяются, за исключением усиления задержки бромсульфалеина. В периферической крови несколько уменьшена масса эритроцитов, уменьшается и объем крови или отмечается транзиторная нормохромная нормоцитарная анемия. Метаболические изменения в период острого приступа заключаются в гиперхолестеринемии на фоне повышения уровня липопротеинов низкой плотности, повышении уровня тироксина в сыворотке (без гипертиреоза), нарушении толерантности к глюкозе и 5а-восстановления тестостерона в печени. Связь этих аномалий с генетическим дефектом остается неясной.
Лечение. Лечение в период острого приступа при ИОП, НКП и ПП одинаково. Некоторые острые приступы можно, по-видимому, купировать путем введения больших количеств (500 г/сут) углеводов (глюкозный эффект), хотя объективных методов исследований эффективности этого лечения не проводилось. Рекомендуется внутривенное введение глюкозы со скоростью 20 г/ч. Если состояние больного за 48 ч непрерывного введения глюкозы не улучшится или если нейропсихическая симптоматика прогрессирует, следует ввести внутривенно гематин (4 мг/кг за 10-15 мин каждые 12 ч в течение 3-6 дней). Он имеется в продаже (пангематин) в виде лиофилизированного порошка. Растворы готовят непосредственно перед инфузией. При использовании гематина в рекомендованных дозах осложнения развиваются крайне редко. Иногда сообщают о тромбофлебите в месте инфузии, коагулопатии (проявляется тромбоцитопенией, удлинением протромбинового времени, некоторым изменением тромбопластинового времени и гипофибриногенемией) и гемолизе. Как гематин, так и глюкоза у экспериментальных животных противодействуют индукции печеночной АЛК-синтазы и за 48 ч могут нивелировать биохимические изменения и обусловить улучшение состояния больного. Для предупреждения и/или коррекции гипонатри-, гипомагни- и азотемии важно проводить поддерживающее лечение при тщательном контроле за состоянием водно-электролитного обмена. Тахикардию и гипертензию следует купировать бета-адреноблокаторами. Если вовремя не установить диагноз при прогрессировании неврологической симптоматики, острые приступы сопряжены с высоким риском смерти. Большинство больных полностью выздоравливают, но неврологическая симптоматика может сохраняться в течение нескольких месяцев и лет. Наиболее важна профилактика острого приступа путем инструктажа больного о необходимости избегать воздействия провоцирующих факторов, например лекарственных веществ, стероидов, потребления алкоголя или намеренного голодания.
T.P. Harrison. Principles of internal medicine. Перевод д.м.н. А. В. Сучкова, к.м.н. Н. Н. Заваденко, к.м.н. Д. Г. Катковского
Порфирии представляют собой патологию, связанную с наследственными или приобретенными аномалиями биосинтеза гема.Порфирины выполняют роль промежуточных продуктов этого пути и образуются из предшественников - d-аминолевулиновой кислоты (АЛК) и порфобилиногена (ПБГ).
Ниже указаны основные этапы синтеза гемма (в скобках указаны ферменты, которые воздействуя на субстрат стоящий с ними в одной строке образуют вещество в ниже идущей строке):
глицин + янтарная кислота – коэнзим А (АЛК-синтетаза)
альфа-амино-бета-кето-адипиновая кислота
дельта-аминолевулиновая кислота (АЛК-дегидрогеназа)
порфобилиноген (ПБГ-дезаминаза)
уропорфириноген lll (Декарбоксилаза)
копропорфириноген lll (КПГ-оксидаза)
протопорфириноген IX (Оксидаза)
протопорфирин IX (Гемсинтетаза)
гем (двухвалентное железо)
глобингемоглобин А
Следует заметить что из порфобилиногена могут образовываться уропорфириноген l и копропорфириноген l при блокировании нормального хода синтеза (значение в развитии порфирии см. далее)
Гем , комплекс двухвалентного железа с протопорфирином IX, функционирует в качестве простетической группы гемопротеинов, таких как гемоглобин, цитохромы, каталаза и триптофаноксигеназа. Его биосинтез жизненно важен и происходит во всех аэробных клетках.
Каждая порфирия характеризуется особенностями гиперпродукции, накопления и экскреции промежуточных продуктов биосинтеза гема. Эти особенности отражают метаболическую экспрессию дефицита отдельных ферментов биосинтеза.
К порфириям не следует относить приобретенные заболевания и интоксикации, при которых выявляется повышенное содержание порфиринов в моче (порфиринурия) или эритроцитах (порфиринемия).
Основные клинические проявления порфирии заключаются в перемежающихся приступах дисфункции нервной системы и/или чувствительности кожи к солнечным лучам.
Неврологический синдром обычно провоцируется приемом таких препаратов, как барбитураты, и заключается в:
болях в животе
периферической нейро¬патии
нарушениях психики
Нейропсихические симптомы появляются только при порфириях, при которых резко усилена продукция предшественников порфиринов - АЛК и порфобилиногена.
Патогенез неврологических нарушений неясен.
Светочувствительность кожи связана непосредственно с повышенным накоплением порфиринов, хотя при разных нарушениях кожные проявления неодинаковы.
Светочувствительность обусловлена фотодинамическим действием порфиринов и опосредуется, вероятно, образующимся синглет-кислородом с последующим развитием деструктивных процессов, таких как перекисное окисление липидов лизосомных мембран.
Доминантно наследуемые порфирии человека экспрессируются по-разному. Могут определяться лишь биохимические или ферментативные изменения. Подобное латентное течение болезни может оказаться одной из стадий или продолжается в течение всей жизни больного. В других случаях симптоматика может провоцироваться лекарственными препаратами, гормонами или повреждением печени.
Классификация
Порфирии подразделяют обычно на две основные группы в соответствии с основными местами синтеза гема, в которых проявляются «ошибки» метаболизма:
эритропоэтическая
печеночная
Единственная чисто эритропоэтическая форма порфирии - врожденная эритро-поэтическая (ВЭП)- встречается редко.
При протопорфирии (ПрП) порфирины накапливаются как в клетках эритроидного ряда, так и в печеночной ткани.
При интермиттирующей острой порфирии, наследственной копропорфирии и пестрой порфирии (соответственно ИОП, НКП и ПП) доминантно наследуемая недостаточность ферментов обусловливает нарушение биосинтеза гема преимущественно в печени без видимых нарушений образования гемоглобина.
Хроническую кожную порфирию (ХКП) ранее считали приобретенной печеночной. Однако у большинства (если не у всех) больных обнаруживают наследственную недостаточность уропорфириногендекарбоксилазы.
Приобретенную порфирию, напоминающую ХКП , обусловливают воздействие полихлорированных углеводородов и опухоли печени.
Отравление свинцом также сопровождается нарушением синтеза порфиринов и гема.
Некоторое усиление экскреции порфиринов или их предшественников с мочой, равно как и накопление порфиринов в эритроцитах, может сопровождать многие клинические состояния.
При вторичных феноменах симптомы и признаки порфирии отсутствуют.
Биохимические аспекты
Последовательность реакций синтеза гема из субстратов глицина и сукцинил-фермента А через АЛК и порфобилиноген (ПБГ) катализируется четырьмя митохондриальными и четырьмя цитозольными ферментами.
В регуляции биосинтеза гема в разных тканях существуют различия. В печени скорость образования гема ограничивается реакцией, катализируемой АЛК-синтазой. Ферменты, действующие после АЛК-синтазы, определяются в избытке. Главным регулятором АЛК-синтазы служит конечный продукт всего пути - гем, который репрессирует фермент по механизму отрицательной обратной связи. Повышенные потребности в геме удовлетворяются путем новообразования АЛК-синтазы. Ее синтез в печени индуцируется большим числом жирорастворимых веществ, стероидами и химическими соединениями, которые служат субстратами и индукторами гемопротеиновых цитохромов P450 - конечных оксидаз на пути микросомного метаболизма фармакологических средств. Эта индукция модулируется многочисленными генетическими, метаболическими и факторами окружа-ющей среды. При порфириях, при которых симптомы провоцируются некоторыми лекарственными препаратами, взаимозависимость синтеза гема и микросомального окисления этих препаратов приобретает большое значение.
В клетках костного мозга, в которых происходит полный синтез гема, ограничивающая скорость реакция также катализируется АЛК-синтазой, но о ее роли в синтезе гема во время деления, дифференцировки и созревания клеток эритроидного ряда известно мало. В процессе созревания этих клеток из них исчезают ядра и митохондрии и, следовательно, митохондриальные ферменты синтеза гема, тогда как цитозольные ферменты, катализирующие реакции между АЛК и копропорфириногеном, сохраняются. В связи с этим эритроциты можно использовать для диагностики порфирий, связанных с дефектом только цитозольного фермента.
Регуляция синтеза гема в костном мозге и печени различна. В печени основной детерминантой образования гема служит уровень АЛК-синтазы, тогда как в костном мозге синтез гема запускается сложным процессом дифференцировки эритроидной клетки. Именно поэтому, вероятно, дефекты ферментов синтеза гема в эритроидных клетках и печени проявляются по-разному.
Порфириногены занимают промежуточное положение между порфобилиногеном и протопорфирином. Они бесцветны и не флюоресцируют. За исключением протопорфирина, порфирины - это побочные продукты, которые покидают путь биосинтеза вследствие необратимого окисления соответствующего порфириногена. Порфирины не выполняют физиологической функции, но в силу своей окраски и флюоресценции определяют необычный цвет мочи и эритроцитов у некоторых больных.
От расположения двух замещенных боковых цепей на пирроловом кольце порфиринов зависят структурные типы изомеров, которые нумеруются от I до IV. В природе найдены только типы I и III, причем только тип III служит субстратом конечных этапов реакции, ведущей к образованию протопорфирина IX и гема. При распаде гема образуются не порфирины, а нециклические тетрапирролы, называемые желчными пигментами.
Врожденная эритропоэтическая порфирия
Врожденная эритропоэтическая порфирия (ВЭП) синонимы:болезнь Гюнтера, врожденная светочувствительная порфирия, эритропоэтическая уропорфирия) - это
редкое
рецессивно наследуемое заболевание
проявляющееся хронической светочувствительностью
проявляющееся обезображивающими повреждениями кожи
проявляющееся гемолитической анемией
Больные представляют собой гомозигот по аутосомно-рецессивному гену. У гетерозигот порфириновый обмен нарушается редко, внешне они выглядят здоровыми.
Патогенез
Лежащий в основе болезни ферментный дефект не установлен, но может заключаться в функциональном неравновесии активностей порфобилиногендезаминазы и косинтазы уропорфириногена III.
Эта аномалия экспрессируется исключительно в созревающих клетках эритроидного ряда и приводит к резкой гиперпродукции уропорфириногена I, тогда как продукция уропорфириногена III не изменяется или несколько повышена. Уропорфириноген I не может использоваться для синтеза гема, но превращается в копропорфириноген I. Уропорфирин I, копропорфириноген I и копропорфирин I накапливаются в тканях и в избыточных количествах экскретируются с мочой и калом.
Клиника
У больных порфирины накапливаются еще в период внутриутробного развития. Уже в родах или вскоре после них обычно начинает отделяться розовая или красная моча, тогда как светочувствительность кожи, периодический гемолиз и спленомегалия могут появиться позднее. Часто определяют гипертрихоз и окрашивание зубов и костей в красный цвет. Смерть может наступить уже в детстве.
При более длительном выживании у больного появляются большие калечащие рубцы, особенно на коже пальцев рук, носа и ушей.
Диагностика
В моче определяют большие количества уропорфирина I, копропорфирина и пор-фиринов с 7, 6,5 и 3 карбоксильными группами, тогда как экскреция АЛК и ПБГ не изменяется.
В кале обнаруживают большие количества копропорфирина I.
Нормобласты, ретикулоциты и эритроциты содержат большие количества уропорфи¬рина I и малое количество копропорфириногена I. Нормобласты и ретикулоциты проявляют интенсивно красную флюоресценцию. В соответствии с нормальной экскрецией АЛ К и ПБГ неврологическая патология отсутствует.
Лечение
Необходимо исключить воздействие солнечного света.
В некоторых случаях гемолитическая анемия, экскреция порфиринов и светочувствительность уменьшаются после спленэктомии.
Применение инфузий гематина и перорального введения b-каротина пока не вышло за рамки эксперимента.
Печеночные порфирии
Три печеночные порфирии (ИОП, НКП и ПП) во многом сходны . Все они наследуются как аутосомный доминантный признак. Острые приступы угрожающей жизни неврологической патологии провоцируются разнообразными препаратами, гормонами и другими факторами, во время которых с мочой экскретируются большие количества АЛК и ПБГ, но виды порфиринов в моче и кале различны.
Интермиттирующая острая порфирия.
Интермиттирующая острая порфирия (ИОП, острая интермиттирующая порфирия (ОИП), пирролопорфирия) характеризуется:
повторными приступами неврологических и психиче¬ских симптомов
светочувствительность отсутствует
первично нарушается функция порфобилиногендезаминазы
Аномалия наследуется как аутосомный доминантный признак с непостоянной экспрессивностью. Аномальный ген встречается с частотой 1:10000-1:50000, но в некоторых регионах она может быть выше. Гомозиготы не встречались.
Патогенез
Причина болезни заключается в частичной (50 %) недостаточности порфобилиногендезаминазы, превращающей ПБГ в уропорфириноген I. На генном уровне эта недостаточность может обусловливаться не одним, а несколькими механизмами, но наиболее частая мутация обусловливает уменьшение количества иммунореактивного фермента белка.
В печени частичная недостаточность фермента приводит к повышению активности и/или индуцибельности АЛК-синтазы лекарственными средствами и другими факторами и, сле-довательно, к повышению образования и экскреции с мочой АЛК и ПБГ. В этих условиях порфирины не накапливаются, и поэтому светочувствительность кожи не повышена.
При ИОП сниженная активность порфобилиногендезаминазы определяется в печени, эритроцитах, культуре кожных фибробластов, лейкоцитах и клетках амниотической жидкости.
Таким образом, ферментный дефект имеет место и во внепеченочных тканях, но его метаболические последствия в них не проявляются. Недостаточность фермента при отсутствии приобретенных факторов необязательно приводит к клинически выраженной острой порфирии, и только у /з больных и даже менее, страдающих этим генетическим дефектом, когда-либо возникает приступ порфирии. Связь между генетическим дефектом и неврологическими нарушениями остается невыясненной.
Клиническая картина
Симптомы болезни редко появляются до пубертатного возраста.
1. Обычно первым и наиболее ярким симптомом приступа порфирии служит боль в животе.
Она может быть умеренной или очень сильной, коликообразной, локализованной или генерализованной, иррадиирует в спину или поясницу.
Боль связана, вероятно, с автономной нейропатией, сопровождающейся нарушением двигательной активности желудочно-кишечного тракта с чередующимися спазматическими и расширившимися участками кишечника.
Живот обычно мягкий, и боль при надавливании не усиливается.
Из-за часто сопутствующих лихорадочного состояния и лейкоцитоза острый приступ порфирии может имитировать любой воспалительный процесс в брюшной полости.
Часты выраженная рвота и запор.
2. Неврологические и психические аномалии проявляются по-разному.
Могут нарушаться функции периферических нервов, автономной нервной системы, ствола мозга, черепных нервов или головного мозга.
Нередки тахикардия и лабильная гипертензия с постуральной гипотензией, задержка мочи и чрезмерная потливость.
Гипертензия и тахикардия коррелируют с повышенной экскрецией катехоламинов.
Периферическая нейропатия обусловливается вовлечением в процесс преимущественно двигательных нервов, но может присоединиться и чувствительный компонент.
Глубокие сухожильные рефлексы снижены или отсутствуют. Типичны невритные боли в конечностях, области гипо- и парестезий, а также вялое свисание стоп и кистей.
Может развиться параплегия или полная вялая квадриплегия.
В прошлом основной причиной смерти служил паралич дыхательной мускулатуры.
При вовлечении в процесс черепных нервов могут атрофироваться зрительный нерв, присоединяться офтальмоплегия и дисфагия.
При более выраженных повреждениях ЦНС появляются бред, кома и судороги.
Несмотря на обратимость нейропатии, остаточные парезы могут сохраняться в течение ряда лет после острого приступа.
Многие больные в течение продолжи¬тельного времени остаются раздражительными, эмоционально неустойчивыми с сохраняющимися функциональными нарушениями.
У 1/3 больных нарушается пси¬хика, могут развиться органический мозговой синдром с беспокойством, дезориентацией и зрительными галлюцинациями.
Иногда определяется выраженная гипонатриемия. Это может быть обусловлено несколькими причинами (в том числе выведением натрия через желудочно-кишечный тракт, неоправданным введением жидкости и сольтеряющей формой нефропатии из-за токсического эффекта АЛК), но ведущей служит, по-видимому, неадекватная секреция антидиуретического гормона.
В ряде случаев присоединяется столь выраженная гипомагниемия, что развивается тетания.
Острые приступы продолжаются в течение нескольких дней и даже месяцев и варьируют по частоте и тяжести. В периоды ремиссий симптомы болезни ослабляются или полностью исчезают.
Клинические и биохимические проявления могут провоцироваться обычными (терапевтические) дозами барбитуратов, противосудо¬рожных средств, эстрогенами, контрацептивами или алкоголем. Все эти вещества окисляются гемопротеинами системы цитохрома Р450. Во время острых приступов метаболизм некоторых из них в печени может нарушаться. У некоторых женщин ухудшение состояния коррелирует с менструальным циклом и латентная порфирия может стать явной в поздние сроки беременности или вскоре после родов. Приступы могут провоцироваться и длительным периодом сниженного потребления калорий (голодание).
Лабораторные данные
Для острых приступов характерна избыточ¬ная экскреция с мочой АЛК и ПБГ, причем по этому признаку ИОП не отличается от НКП или ПП. Уровень АЛК и ПБГ в моче не коррелирует с выраженностью симптоматики. Простым и надежным скрининг-тестом, помогающим диагностировать острый приступ ИОП, НКП и ПП, служит качественное определение порфобилиногена в моче (тесты Уотсона -Шварца или Хоша). При нейропсихических нарушениях эти тесты почти всегда положительны, однако для этого требуется, чтобы концентрация ПБГ в моче превышала верхнюю границу нормы в 3-5 раз. В связи с этим оба теста при латентной форме болезни или нормализации экскреции ПБГ после купирования приступа могут быть отрицательными. Иногда требуется количественное определение экскретируемых с мочой АЛК и ПБГ с помощью хроматографических методов.
При латентной форме ИОП с нормальной экскрецией АЛК и ПБГ диагноз может быть установлен на основании результатов определения активности порфобилиногендезаминазы в эритроцитах, лейкоцитах или культивируемых фибробластах кожи. Однако у здоровых и больных ИОП эти результаты перекрываются, и точно поставить диагноз не всегда возможно.
При ИОП в соответствии с ферментным дефектом усилена экскреция пред-шественников порфиринов - АЛК и ПБГ, поэтому свежеполученная моча обычно бесцветна, содержит мало предобразованного уро- или копропорфирина. При стоянии она может темнеть, так как ПБГ спонтанно полимеризуется в уропорфирин и порфобилин - темно-коричневый пигмент неизвестного строения. У некоторых больных, однако, определяют достаточное количество образующихся нефермента-тивным путем пигментов, чтобы придавать свежеполученной моче темно-красный цвет.
Концентрация порфиринов в кале обычно находится в пределах нормы.
Общепринятые функциональные печеночные пробы не изменяются, за исклю-чением усиления задержки бромсульфалеина.
В периферической крови несколько уменьшена масса эритроцитов, уменьшается и объем крови или отмечается транзиторная нормохромная нормоцитарная анемия.
Метаболические изменения в период острого приступа заключаются в гиперхолестеринемии на фоне повышения уровня липопротеинов низкой плотности, повышении уровня тироксина в сыворотке (без гипертиреоза), нарушении толерантности к глюкозе и 5а-восстановления тестостерона в печени. Связь этих аномалий с генетическим дефектом остается неясной.
Лечение
Лечение в период острого приступа при ИОП, НКП и ПП одинаково.
Некоторые острые приступы можно, по-видимому, купировать путем введения больших количеств (500 г/сут) углеводов (глюкозный эффект), хотя объек¬тивных методов исследований эффективности этого лечения не проводилось.
Реко¬мендуется внутривенное введение глюкозы со скоростью 20 г/ч. Если состояние больного за 48 ч непрерывного введения глюкозы не улучшится или если нейропсихическая симптоматика прогрессирует, следует ввести внутривенно гематин (4 мг/кг за 10-15 мин каждые 12 ч в течение 3-6 дней). Он имеется в продаже (пангематин) в виде лиофилизированного порошка. Растворы готовят непосредственно перед инфузией. При использовании гематина в рекомендованных дозах осложнения развиваются крайне редко. Иногда сообщают о тромбофлебите в месте инфузии, коагулопатии (проявляется тромбоцитопенией, удлинением протромбинового времени, некоторым изменением тромбопластинового времени и гипофибриногенемией) и гемолизе.
Как гематин, так и глюкоза у экспериментальных животных противодействуют индукции печеночной АЛК-синтазы и за 48 ч могут нивелировать биохимические изменения и обусловить улучшение состояния больного.
Для предупреждения и/или коррекции гипонатри-, гипомагни- и азотемии важно проводить поддерживающее лечение при тщательном контроле за состоянием водно-электролитного обмена.
Тахикардию и гипертензию следует купировать бета-адреноблокаторами.
Если вовремя не установить диагноз при прогрессировании неврологической симп-томатики, острые приступы сопряжены с высоким риском смерти.
Большинство больных полностью выздоравливают, но неврологическая симптоматика может сохраняться в течение нескольких месяцев и лет.
Наиболее важна профилактика острого приступа путем инструктажа больного о необходимости избегать воздействия провоцирующих факторов, например лекарственных веществ, стероидов, потребления алкоголя или намеренного голодания.
Наследственная копропорфирия
Наследственная копропорфирия (НКП) представляет собой печеночную порфирию, характеризующуюся приступами нейропсихических расстройств, идентичных таковым при ИОП и ПП.
У некоторых больных повышена светочувствительность кожи.
Первичный генетический дефект заключается в частичной недостаточности копропорфириногеноксидазы.
Болезнь наследуется как аутосомный доминантный признак.
Частота ее неизвестна, так как в большинстве случаев клиническая симптоматика отсутствует.
При НКП экскретируются большие количества копропорфирина III, особенно с калом.
Экскреция АЛК и ПБГ усиливается во время острых приступов (положительные тесты Уотсона -Шварца или Хоша), но в период ремиссии она обычно остается в пределах нормы. Острые приступы неотличимы от таковых при ИОП и ПП и провоцируются теми же факторами.
Светочувствительность кожи повышена примерно у 1/3 больных. Частичную недостаточность копропорфириногеноксидазы можно обнаружить в лейкоцитах и культуре кожных фибробластов.
Лечение идентично таковому при ИОП.
Пестрая порфирия
Пестрая порфирия (ПП, южноафриканская генетическая порфирия) характеризуется как острыми приступами нейропсихических нарушений, так и хронической чувствительностью кожи к солнечному свету и механическим травмам.
Первичный ферментный дефект на пути синтеза гема заключается
в частичной недостаточности протопорфириногеноксидазы.
Пестрая порфирия наследуется как аутосомный доминантный признак.
Она особенно распространена среди представителей европеоидной популяции Южной Африки, где ее частоту оценивают в 1:400. В других регионах больные встречаются реже, но все же их выявляют во многих странах.
Ферментный дефект обусловливает экскрецию больших количеств протопорфирина с желчью и калом (с менее выраженным усилением экскре¬ции копропорфирина с калом) и АЛК, ПБГ и копропорфирина с мочой во время острых приступов.
Явная ПП развивается обычно на втором-третьем 10-летии жизни.
Клиника
заключается в острых приступах болей в животе и нейропсихических нарушениях на фоне повреждения кожи, вызванного солнечными лучами.
Неврологические и кожные проявления могут сосуществовать или появляются в разное время. У большинства больных, проживающих в Южной Африке, после незначительных механических травм на открытых участках кожи появляются ссадины, поверхностные эрозии и волдыри. На их месте часто остаются депигментированные или пигментированные рубцы. Заживление задерживается при вторичной инфекции. Кожа лица и рук у больных обычно гиперпигментирована, а женщины часто страдают гирсутизмом. Кожные повреждения неотличимы от таковых при хронической кожной порфирии (ХКП). Резкое обострение кожных изменений может быть связано со случайными заболеваниями печени, предположительно сопровождающимися снижением экскреции порфиринов с калом и одновременно усилением их экскреции с мочой.
Острые приступы нейропсихических расстройств неотличимы от таковых при ИОП и НКП и провоцируются теми же факторами.
Характерным химическим признаком служит постоянная экскреция больших количеств прото- и копропорфирина даже в тех случаях, когда клиническая симптоматика минимальна или вообще отсутствует. Количество протопорфирина превышает количество копропорфирина, т. е. ситуация обратна той, которая имеет место при НКП. Экскреция АЛК, ПБГ и порфиринов с мочой у больных при бессимптомной форме или у тех, у кого появляются только кожные изменения, либо не изменяется, либо несколько усилена.
Во время острых приступов экскреция АЛК и ПБГ с мочой повышается (положительные тесты Уотсона - Шварца или Хоша), увеличивается и экскреция с мочой копропорфирина и уропорфирина. Содержание порфиринов в эритроцитах находится в пределах нормы, что отличает пеструю порфирию от протопорфирии.
Лечение - профилактика и лечение при острых приступах инфузиями глюкозы и, возможно, гематина те же, что и при ИОП и НКП, хотя опыт применения гематина при ПП более ограничен. Рекомендуется избегать воздействия прямых солнечных лучей и носить защитную одежду (шляпы, перчатки). Прогноз сходен с таковым при ИОП или несколько более благоприятен.
Хроническая кожная порфирия
Хроническая кожная порфирия (ХКП, симптоматическая кожно-печеночная порфирия, симптоматическая порфирия) - наиболее распространенная из всех порфирий.
Она характеризуется хроническими изменениями кожи, частой печеночной патологией (и печеночным сидерозом), особенностью экскреции порфиринов с мочой.
Заболевание обусловлено, вероятно, врожденной или приобретенной недостаточностью печеночной уропорфириногендекарбоксилазы.
Неврологические нарушения отсутствуют.
Хроническую кожную порфирию считали приобретенным заболеванием из-за ее спорадического (и обычно несемейного характера) начала в зрелом возрасте и частой связи с алкогольным повреждением печени и ее сидерозом.
Распространенность болезни не установлена, но у больных алкоголизмом с перегрузкой железом развивается часто, как, например, среди племен банту в Южной Африке.
Она может быть и семейной патологией , передаваясь как аутосомный доминантный признак с непостоянной экспрессивностью (семейная ХКП).
Врожденный дефект заключается в некотором снижении активности уропорфириноген-декарбоксилазы в печени, эритроцитах и культуре фибробластов. Клинически и химически выявлены латентные носители дефекта.
При спорадической ХКП частичная недостаточность уропорфириногендекарбоксилазы обнаруживается только в печени. Неизвестно, является ли это результатом генетического или приобретенного (токсический) механизма. Недостаточность (какова бы ни была ее этиология) уропорфириногендекарбоксилазы, которая катализирует превращение уро-порфириногена в копропорфириноген, приводит к нарушению печеночного синтеза гема с последующим развитием светочувствительности кожи только при воздействии дополнительных факторов, например перегрузки железом, обычно в сочетании с повреждением печени и длительным приемом эстрогенов.
Механизм, посредством которого перегрузка железом и гормоны вызывают клиническую экспрессию латентной ХКП, неизвестен.
В отличие от ИОП, НКП и ПП ферментный дефект при ХКП не служит результатом нарушенной регуляции в цепи реакций синтеза гема в печени, и активность АЛК-синтазы не изменяется или лишь несколько усиливается даже при явной патологии. Это, вероятно, объясняет отсутствие острых нейропсихических приступов, обычно нормальный уровень АЛК и ПБГ в моче и переносимость таких средств, как барбитураты.
Клинические проявления - единственным выраженным проявлением болезни служит повышенная светочувствительность кожи. Ее изменения идентичны таковым при ПП. Они обычно появляются исподволь, чаще всего у мужчин в возрасте 40-60 лет, и заключаются в усиленной пигментации кожи лица, повышении ее чувствительности к травмам, в эритеме, а также появлении пузырей и язв. Часто появляются склеродерматозные изменения и чрезмерный рост волос в области лба, скуловой области и предплечий.
Обычно выявляют патологию печени, часто связанную с алкоголизмом, и почти всегда - ее сидероз, хотя количество скопившегося железа варьирует и редко бывает значительным. Может наступить спонтанная ремиссия.
Иногда клиническая симптоматика провоцируется эстрогенами (включая контрацептивы) или известными гепатотоксическими веществами. При ХКП увеличена частота сахарного диабета, кроме того, она сочетается с системной красной волчанкой и другими аутоиммунными синдромами.
Экскреция уропорфирина и в меньшей степени копропорфирина с мочой усиливается. Моча может иметь розовый или коричневый цвет. Экскреция АЛК и ПБГ с мочой обычно не изменяется (отрицательные тесты Уотсона-Шварца или Хоша). Кроме уропорфирина, основного порфирина мочи, в ней находят и промежуточные порфирины (в частности, гептакарбоксильный). Увеличение содержания порфиринов в кале выражено слабее и обычно ограничено копропорфириновой фракцией.
Диагноз устанавливают на основании сочетания повышения светочувствительности кожи, печеночной патологии, повышенной экскреции уропорфирина с мочой, отсутствия увеличения предшественников порфиринов (АЛК, ПБГ) и нейропсихических приступов в анамнезе.
Приобретенная токсическая порфирия, напоминающая ХКП, может развиться у лиц, случайно подвергшихся воздействию гексахлорбензена, полихлорированных бифенилов, тетрахлордибензо-р-диоксина (ТХДД) и других полихлорированных углеводородов. Кроме того, известно несколько случаев ХКП, сочетающейся с доброкачественными или злокачественными первичными опухолями печени.
Лечение . Отказ от приема алкоголя обычно приводит к улучшению состояния больного. Выведение железа из печени путем повторных кровопусканий также может обусловливать длительную ремиссию: еженедельно или реже забирают 400 мл крови (или эквивалентное количество эритроцитов) при тщательном наблюдении за уровнем гемоглобина и белков плазмы. У больных, которым кровопускание противопоказано, вывести уропорфирины из печени и добиться ремиссии можно, вероятно, с помощью малых доз хлорохина (125 мг дважды в неделю). Однако он может оказывать токсическое действие на печень. Альтернативой служит также хелатообразующее средство дефероксамин. Местное экранирование от солнца и пероральный прием каротиноидов не оказывают защитного действия.
Протопорфирии
Протопорфирия (ПрП, эритропоэтическая протопорфирия, эритропеченочная протопорфирия)- заболевание, при котором некоторая светочувствительность кожи сочетается с высокой концентрацией протопорфирина в эритроцитах, обусловленной недостаточностью феррохелатазы. Протопорфирин может накапливаться и в печени.
Протопорфирия наследуется как аутосомный доминантный признак с непостоянной экспрессивностью.
Активность феррохелатазы, катализирующей включение двухвалентного железа в протопорфирин, снижена в костном мозге, периферической крови, печени и культивируемых фибробластах кожи. Эта недостаточность приводит к чрезмерному накоплению протопорфирина в зрелых нормобластах, ретикулоцитах и молодых эритроцитах. По мере старения эритроцитов протопорфирин поступает из них в плазму.
Светочувствительность кожи опосредуется протопорфирином плазмы и кожи и вызывается видимой частью спектра (380-560 нм). Светочувствительность кожи обнаруживает сезонную вариабельность.
У некоторых больных печень принимает участие в избыточной продукции порфиринов или, наоборот, может поглощать протопорфирин из плазмы.
Многие носители этого дефекта остаются клинически (и химически) здоровыми, и диагноз можно установить только с помощью исследования ферментов.
Клинические проявления
. Некоторая светочувствительность кожи возникает обычно в детстве. Пребывание на солнце приводит к появлению зуда, эритемы и иногда отека (солнечная крапивница). Через несколько часов или дней эти явления стихают, не оставляя рубцов. Кожные проявления могут возникать лишь после длительного пребывания на солнце. В других случаях начальные изменения кожи прогрессируют до хронической экзематозной фазы (солнечная экзема). При этом заболевании кожа достаточно устойчива к механическим воздействиям, на ней не образуются волдыри в отличие от ПП и ХКП. Не определяются также эритродонтия, гипертрихоз и гиперпигментация. Не наступает и приступов нейропсихических нарушений.
Протопорфирия - болезнь обычно доброкачественная, но может сопровождаться патологией печени, желчных путей или крови. При ней увеличивается частота холелитиаза, причем в состав желчных камней входит протопорфирин. Иногда патология печени вследствие отложения большого количества протопорфирина может прогрессировать до цирроза, приводящего к смерти больного, поэтому у каждого больного следует проводить обычные функциональные печеночные пробы. Нередко протопорфирия сопровождается некоторой анемией.
Диагноз основан на обнаружении высоких концентраций протопорфирина в эритроцитах. При флюоресцентной микроскопии можно видеть большое число флюоресцирующих красным цветом эритроцитов. Уровень протопорфирина может быть повышен и в плазме и кале, тогда как в моче его количество, а также АЛК и ПБГ обычно не изменяется.
Лечение . Местная защита от солнца обычно неэффективна. Пероральный прием b-каротина (обычно в виде смеси р-каротина и кантаксантина) существенно повышает переносимость солнечных лучей. Содержание каротина в сыворотке следует поддерживать на уровне 6000-8000 мкг/л.





