Бактерия жасушаларының өсуі мен көбеюінің ерекшелігі неде. Бактериялардың тіршілік ету және өсу фазалары Микроорганизмдердің көбею кезеңдері
Өсу және көбею
«Өсу» термині жасушалық материалдың (мысалы, ақуыз, РНҚ, ДНҚ) синтезі нәтижесінде жеке жасушаның немесе бактериялар тобының цитоплазмалық массасының ұлғаюын білдіреді. Белгілі бір мөлшерге жеткеннен кейін жасуша өсуі тоқтап, көбейе бастайды.
Микробтардың көбеюі олардың өздігінен көбею қабілетін, көлем бірлігіне даралардың санын көбейтуді білдіреді. Әйтпесе, айта аламыз: көбею - микроб популяциясының дараларының көбеюі.
Бактериялар негізінен қарапайым көлденең бөліну (вегетативті көбею) арқылы көбейеді, ол әртүрлі жазықтықта, әр түрлі жасушалар комбинациясын (жүзім шоғы – стафилококк, тізбек – стрептококк, жұп – диплококк, бума, бума – сарцин және т.б.) түзе отырып жүреді. Бөлу процесі бірнеше кезекті кезеңдерден тұрады. Бірінші кезең жасушаның ортаңғы бөлігінде көлденең қалқаның пайда болуымен басталады (6-сурет), ол бастапқыда аналық жасушаның цитоплазмасын екі еншілес жасушаға бөлетін цитоплазмалық мембранадан тұрады. Осымен қатар жасуша қабырғасы синтезделеді, ол екі еншілес жасушалар арасында толыққанды бөлімді құрайды. Бактериялардың бөліну процесінде ДНҚ-ның репликациясы (екі еселенуі) маңызды шарт болып табылады, оны ДНҚ-полимераза ферменттері жүзеге асырады. ДНҚ екі еселенгенде сутектік байланыстар үзіліп, ДНҚ-ның екі тізбегі түзіледі, олардың әрқайсысы еншілес жасушаларда орналасады. Әрі қарай қызы бір тізбекті ДНҚ сутегі байланыстарын қалпына келтіреді және қайтадан қос тізбекті ДНҚ құрайды.
ДНҚ репликациясы мен жасушаның бөлінуі микробтардың әрбір түріне тән белгілі бір жылдамдықпен жүреді, ол дақылдың жасына және қоректік ортаның табиғатына байланысты. Мысалы, ішек таяқшасының өсу жылдамдығы 16 минуттан 20 минутқа дейін; туберкулез микобактериясында бөліну 18-20 сағаттан кейін ғана жүреді; сүтқоректілердің тіндік культурасының жасушасы 24 сағатты алады. Демек, көптеген түрлердің бактериялары тіндік жасушаларға қарағанда 100 есе жылдам көбейеді.
Ауыстырылмайтын ортада микробтық дақылдың көбею процесі біркелкі емес жүреді. Ол төрт негізгі кезеңді анықтайды.
1. Бастапқы кезең (кешіктірілген фаза) немесе тыныштық кезеңі.Бұл уақытта дақыл қоректік ортаға бейімделеді. Микроб жасушасында РНҚ мөлшері артып, оның көмегімен қажетті ферменттердің синтезі жүреді.
2. Көрсеткіштік (логарифмдік) фазакультурадағы жасушалардың максималды ұлғаюымен сипатталады, ол экспоненциалды түрде өтеді (1, 2.4, 8, 16, 256 және т.б.). Бұл кезде жас және биологиялық белсенді жасушалардың көпшілігі ортада болады. Фазаның соңында, орта таусылғанда, берілген микробқа қажетті заттар жоғалады, оттегінің мөлшері азаяды, зат алмасу өнімдерінің көбеюі орын алады - культураның өсуі баяулайды. Қисық бірте-бірте көлденең бағытты алады.
3. стационарлық фаза,немесе өтеу кезеңі графикалық түрде x осіне параллель болатын сызықты көрсетеді. Жаңадан пайда болған және өлі жасушалардың саны арасында тепе-теңдік бар. Ортаның мөлшері азаяды, популяциядағы жасушалардың тығыздығы артады, метаболизм өнімдерінің токсикалық әсері күшейеді - мұның бәрі жасушалардың өліміне әкеледі.
4. Өлу кезеңі.Бұл фазада тек азайып қана қоймай, жасушалардың өзгеруі де байқалады. Деградацияланған формалар, сондай-ақ споралар пайда болады. Бірнеше аптадан немесе айдан кейін мәдениет өледі. Бұл улы қалдықтардың микроб жасушаларын тежеп қана қоймай, сонымен бірге өлтіруімен байланысты.
Осылайша, метаболизм процестерінің арқасында микроб жасушасының өмірлік белсенділігі сақталады. Аэробтар тыныс алу үшін оттегіні қажет етеді, ал анаэробтар нитраттық және сульфатты тыныс алуды және ашытуды пайдаланады. Микроорганизмдер сыртқы ортадан органикалық және бейорганикалық заттарды ассимиляциялайды, олар тотықтырады, олар қажетті энергия мен пластикалық элементтерді алады. Нәтиже - жасушаның өсуі. Қажетті жетілу кезеңіне жеткен жасуша қарапайым бөліну арқылы көбейеді. Микроорганизмдер тіршілік әрекеті барысында қоректік заттарды бірте-бірте тұтынады, олардың метаболиттерін қоршаған ортаға шығарады, сол арқылы қоршаған ортаның құрамын өзгертіп, тіршілікке жарамсыз етеді.
Бактериялардың бөліну арқылы көбеюі микробтар популяциясының мөлшерін көбейтудің ең кең таралған әдісі болып табылады. Бөлінгеннен кейін бактериялар бастапқы мөлшеріне дейін өседі, бұл белгілі бір заттарды (өсу факторларын) қажет етеді.
Бактериялардың көбею әдістері әртүрлі, бірақ олардың көптеген түрлері үшін жыныссыз көбею формасы бөлу әдісіне тән. Бактериялар бүршіктену арқылы сирек көбейеді. Бактериялардың жыныстық көбеюі қарабайыр формада болады.
Күріш. 1. Фотосуретте бактерия жасушасы бөліну сатысында.
Бактериялардың генетикалық аппараты
Бактериялардың генетикалық аппараты бір ДНҚ – хромосомамен ұсынылған. ДНҚ сақинада тұйықталған. Хромосома мембранасы жоқ нуклеотидте орналасқан. Бактерия жасушасында плазмидалар болады.
Нуклеоид
Нуклеоид ядроға ұқсас. Ол жасушаның ортасында орналасқан. ДНҚ онда локализацияланған - тұқым қуалайтын ақпаратты бүктелген түрде тасымалдаушы. Бұралмаған ДНҚ ұзындығы 1 мм-ге жетеді. Бактерия жасушасының ядролық затында қабықша, ядрошық және хромосомалар жиынтығы болмайды, митоз жолымен бөлінбейді. Бөліну алдында нуклеотид екі еселенеді. Бөліну кезінде нуклеотидтер саны 4-ке дейін артады.

Күріш. 2. Суретте кесілген жердегі бактерия жасушасы. Орталық бөлігінде нуклеотид көрінеді.
Плазмидалар
Плазмидалар – қос тізбекті ДНҚ сақинасына бүктелген автономды молекулалар. Олардың массасы нуклеотидтің массасынан әлдеқайда аз. Тұқым қуалайтын ақпарат плазмидалардың ДНҚ-да кодталғанына қарамастан, олар бактериялық жасуша үшін өмірлік маңызды және қажет емес.

Күріш. 3. Фотода бактериялық плазмида көрсетілген.
Бөлу кезеңдері
Ересек жасушаға тән белгілі бір мөлшерге жеткеннен кейін бөліну механизмдері іске қосылады.
ДНҚ репликациясы
ДНҚ репликациясы жасушаның бөлінуінен бұрын жүреді. Мезосомалар (цитоплазмалық мембрананың қатпарлары) ДНҚ-ны бөліну (репликация) процесі аяқталғанша ұстайды.
ДНҚ репликациясы ДНҚ-полимераза ферменттерінің көмегімен жүзеге асады. Репликация кезінде 2 тізбекті ДНҚ-дағы сутектік байланыстар үзіледі, нәтижесінде бір ДНҚ-дан екі қызы бір тізбекті байланыстар түзіледі. Кейінірек, бөлінген аналық жасушаларда қыз ДНҚ орын алған кезде, олар қалпына келтіріледі.
ДНҚ репликациясы аяқталғаннан кейін синтез нәтижесінде жасушаны екіге бөлетін тарылу пайда болады. Алдымен нуклеотид бөлінеді, содан кейін цитоплазма. Жасуша қабырғасының синтезі бөлінуді аяқтайды.
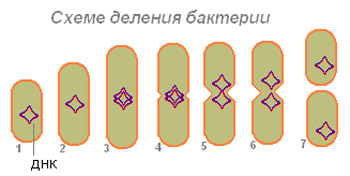
Күріш. 4. Бактерия жасушасының бөліну схемасы.
ДНҚ сегменттерінің алмасуы
Шөп таяқшасында ДНҚ репликация процесі 2 ДНҚ сегментінің алмасуымен аяқталады.
Жасушаның бөлінуінен кейін көпір пайда болады, оның бойымен бір жасушаның ДНҚ-сы екіншісіне өтеді. Содан кейін екі ДНҚ өзара тоғысады. Екі ДНҚ-ның кейбір бөліктері бір-біріне жабысады. Адгезия орындарында ДНҚ сегменттері алмасады. ДНҚ-ның бірі секіргіш бойындағы бірінші ұяшыққа қайта оралады.

Күріш. 5. Пішен таяқшасындағы ДНҚ алмасу нұсқасы.
Бактерия жасушасының бөліну түрлері
Егер жасушаның бөлінуі бөліну процесінен озып кетсе, онда көп жасушалы таяқшалар мен кокктар түзіледі.
Синхронды жасушалардың бөлінуімен екі толыққанды еншілес жасушалар пайда болады.
Егер нуклеотид жасушаның өзінен жылдам бөлінсе, онда мультинуклеотидті бактериялар түзіледі.
Бактерияларды бөлу жолдары
Бөлу арқылы бөлу
Сібір жарасының таяқшаларына ыдырату арқылы бөліну тән. Осы бөліну нәтижесінде жасушалар буындарда үзіліп, цитоплазмалық көпірлер бұзылады. Содан кейін олар бір-бірін итермелейді, тізбектер жасайды.
сырғымалы бөлу
Бөлінгеннен кейін жылжымалы бөліну арқылы жасуша бөлінеді және басқа жасушаның бетімен сырғанайды. Бұл бөлу әдісі эшерихияның кейбір түрлеріне тән.
бөлу бөлу
Бөлінген бөліну кезінде бөлінген жасушалардың бірі бос ұшы бар шеңбер доғасын сипаттайды, оның орталығы оның басқа жасушамен жанасу нүктесі болып табылады, римдік бес немесе сына тәрізді (коринебактерия дифтерия, листерия) құрайды.

Күріш. 6. Фотосуретте таяқша тәрізді бактериялар тізбек түзеді (сібір жарасы таяқшалары).

Күріш. 7. Фотосуретте ішек таяқшасын бөлудің сырғымалы әдісі.

Күріш. 8. Коринебактерияларды бөлу әдісі.
Бөлінгеннен кейінгі бактериялық кластерлердің көрінісі
Бөлінетін жасушалардың жинақталуы бөліну жазықтығының бағытына байланысты әртүрлі пішіндерге ие.
глобулярлы бактерияларбір-бірден, екіден (диплококктар), сөмкелерде, шынжырларда немесе жүзім шоқтары сияқты орналасады. Таяқша тәрізді бактериялар – тізбекте.
Спиральды бактериялар- хаотикалық.

Күріш. 9. Фотосуретте микрококктар көрсетілген. Олар дөңгелек, тегіс, ақ, сары және қызыл түсті. Микрококктар табиғатта барлық жерде кездеседі. Олар адам денесінің әртүрлі қуыстарында өмір сүреді.

Күріш. 10. Суретте диплококк бактериялары - Streptococcus pneumoniae.

Күріш. 11. Фотодағы сарцина бактериялары. Коккоидты бактериялар пакеттерге біріктіріледі.

Күріш. 12. Фотосуретте стрептококк бактериялары (грек тілінен аударғанда «стрептос» - тізбек). Тізбекпен реттелген. Олар бірқатар аурулардың қоздырғыштары болып табылады.

Күріш. 13. Фотосуретте бактериялар «алтын» стафилококктар. «Жүзім шоғыры» сияқты реттелген. Кластерлердің алтын түсі бар. Олар бірқатар аурулардың қоздырғыштары болып табылады.

Күріш. 14. Фотосуретте лептоспиралардың конвульсиялы бактериялары көптеген аурулардың қоздырғыштары болып табылады.

Күріш. 15. Суретте Vibrio тұқымдасының таяқша тәрізді бактериялары.
бактериялардың бөліну жылдамдығы
Бактериялардың бөліну жылдамдығы өте жоғары. Орташа алғанда бір бактерия жасушасы әрбір 20 минут сайын бөлінеді. Бір күннің ішінде бір жасуша ұрпақтың 72 ұрпағын құрайды. Туберкулез микобактериясы баяу бөлінеді. Бүкіл бөлу процесі оларға шамамен 14 сағатты алады.

Күріш. 16. Фотода стрептококк жасушасының бөліну процесі көрсетілген.
Бактериялардың жыныстық көбеюі
1946 жылы ғалымдар қарабайыр формада жыныстық көбеюді ашты. Бұл жағдайда гаметалар (еркек және аналық жыныс жасушалары) түзілмейді, бірақ кейбір жасушалар генетикалық материалмен алмасады ( генетикалық рекомбинация).
нәтижесінде геннің тасымалдануы жүреді конъюгациялар— түрдегі генетикалық ақпараттың бір бөлігін бір бағытты беру плазмидабактериялық жасушалар арасындағы байланыста.
Плазмидалар – кішігірім ДНҚ молекулалары. Олар хромосома геномымен байланысты емес және автономды түрде қайталануға қабілетті. Плазмидаларда бактерия жасушаларының қоршаған ортаның қолайсыз жағдайларына төзімділігін арттыратын гендер бар. Бактериялар көбінесе бұл гендерді бір-біріне береді. Басқа түрдің бактерияларына гендік ақпараттың берілуі де атап өтіледі.
Шынайы жыныстық процесс болмаған жағдайда, пайдалы қасиеттердің алмасуында үлкен рөл атқаратын конъюгация. Бұл бактериялардың дәріге төзімділік көрсету қабілетін береді. Адамзат үшін ауру тудыратын популяциялар арасында антибиотиктерге төзімділіктің берілуі ерекше қауіпті.

Күріш. 17. Суретте екі ішек таяқшасының конъюгациясы сәті.
Бактериялар популяциясының даму фазалары
Қоректік ортаға себу кезінде бактерия популяциясының дамуы бірнеше фазалардан өтеді.
Бастапқы кезең
Бастапқы кезең - себу сәтінен бастап олардың өсуіне дейінгі кезең. Орташа алғанда, бастапқы кезең 1-2 сағатқа созылады.
Репродуктивті кідіріс фазасы
Бұл бактериялардың қарқынды өсу кезеңі. Оның ұзақтығы шамамен 2 сағатты құрайды. Ол дақылдың жасына, бейімделу кезеңіне, қоректік ортаның сапасына және т.б.
логарифмдік фаза
Бұл фазада бактериялардың көбею жылдамдығының және көбеюінің шыңы байқалады. Оның ұзақтығы 5-6 сағатты құрайды.
Теріс үдеу фазасы
Бұл фазада көбею жылдамдығының төмендеуі байқалады, бөлінетін бактериялардың саны азайып, өлі бактериялардың саны артады. Теріс жеделдетудің себебі - қоректік ортаның таусылуы. Оның ұзақтығы шамамен 2 сағатты құрайды.
Стационарлық максимум фаза
Стационарлық фазада өлген және жаңадан пайда болған особьтардың тең саны белгіленеді. Оның ұзақтығы шамамен 2 сағатты құрайды.
Жеделдетілген өлім фазасы
Бұл кезеңде өлі жасушалардың саны біртіндеп артады. Оның ұзақтығы шамамен 3 сағатты құрайды.
Логарифмдік өлім фазасы
Бұл фазада бактерия жасушалары тұрақты жылдамдықпен өледі. Оның ұзақтығы шамамен 5 сағатты құрайды.
Төмендеу фазасы
Бұл фазада қалған тірі бактерия жасушалары тыныштық күйіне өтеді.

Күріш. 18. Суретте бактерия популяциясының өсу қисығы көрсетілген.

Күріш. 19. Фотосуретте Pseudomonas aeruginosa-ның көк-жасыл колониялары, микрококктардың сары колониялары, Bacterium prodigiosum-ның қанды-қызыл колониялары және Bacteroides niger қара колониялары көрсетілген.

Күріш. 20. Фотода бактериялар колониясы көрсетілген. Әрбір колония бір жасушаның ұрпағы болып табылады. Колонияда жасушалардың саны миллиондаған. колония 1-3 күнде өседі.
Магниттік сезімтал бактериялардың бөлінуі
1970 жылдары теңіздерде тіршілік ететін магниттік сезімі бар бактериялар табылды. Магнитизм бұл таңғажайып жаратылыстарға Жердің магнит өрісінің сызықтары бойынша жүруге және оған қажетті күкірт, оттегі және басқа заттарды табуға мүмкіндік береді. Олардың «компастары» магниттен тұратын магнитосомалармен бейнеленген. Бөлу кезінде магнитті сезімтал бактериялар компасын бөледі. Бұл жағдайда бөліну кезіндегі тарылту айқын жеткіліксіз болады, сондықтан бактерия жасушасы иіліп, өткір сынық жасайды.

Күріш. 21. Фотосуретте магнитті сезімтал бактерияның бөліну сәті көрсетілген.
Бактериялардың көбеюі
Бактерия жасушасының бөлінуінің басында екі ДНҚ молекуласы жасушаның әртүрлі ұштарына бөлінеді. Әрі қарай ұяшық екі тең бөлікке бөлінеді, олар бір-бірінен бөлініп, бастапқы өлшеміне дейін ұлғаяды. Көптеген бактериялардың бөліну жылдамдығы орта есеппен 20-30 минутты құрайды. Бір күннің ішінде бір жасуша ұрпақтың 72 ұрпағын құрайды.
Өсу және даму процесіндегі жасушалардың массасы қоршаған ортадан қоректік заттарды тез сіңіреді. Бұған қолайлы қоршаған орта факторлары - температура, қоректік заттардың жеткілікті мөлшері, қоршаған ортаның қажетті рН-ы ықпал етеді. Аэробты жасушалар оттегін қажет етеді. Анаэробтар үшін бұл қауіпті. Алайда, табиғатта бактериялардың шексіз көбеюі болмайды. Бактерия жасушасына күн сәулесі, құрғақ ауа, тағамның жетіспеушілігі, қоршаған ортаның жоғары температурасы және басқа факторлар зиянды әсер етеді.

Күріш. 22. Фотосуретте жасушаның бөліну сәті.
өсу факторлары
Бактериялардың көбеюі үшін белгілі бір заттар (өсу факторлары) қажет, олардың кейбіреулері жасушаның өзінде синтезделеді, ал кейбіреулері сыртқы ортадан келеді. Барлық бактериялардың өсу факторларына әртүрлі талаптары бар.
Өсу факторларының қажеттілігі тұрақты қасиет болып табылады, бұл оны бактерияларды анықтау, қоректік орталарды дайындау және биотехнологияда қолдану үшін пайдалануға мүмкіндік береді.
Бактериялардың өсу факторлары (бактериялық витаминдер) химиялық элементтер, олардың көпшілігі суда еритін В тобының витаминдері.Бұл топқа сонымен қатар гемин, холин, пурин және пиримидин негіздері және басқа да аминқышқылдары жатады. Өсу факторлары болмаған жағдайда бактериостаз пайда болады.
Бактериялар өсу факторларын ең аз мөлшерде және өзгеріссіз пайдаланады. Бұл топтағы бірқатар химиялық заттар жасушалық ферменттердің бөлігі болып табылады.

Күріш. 23. Суретте таяқша тәрізді бактерияның бөліну сәті.
Бактериялардың өсуінің маңызды факторлары
- В1 дәрумені (тиамин). Көмірсулар алмасуына қатысады.
- В2 дәрумені (рибофлавин). Тотығу-тотықсыздану реакцияларына қатысады.
- Пантотен қышқылыкофермент А-ның құрамдас бөлігі болып табылады.
- В6 дәрумені (пиридоксин). Амин қышқылдарының алмасуына қатысады.
- В12 витаминдері(кобаламиндер – құрамында кобальт бар заттар). Олар нуклеотидтердің синтезіне белсенді қатысады.
- Фолий қышқылы. Оның кейбір туындылары пуриндік және пиримидиндік негіздердің, сондай-ақ кейбір аминқышқылдарының синтезін катализдейтін ферменттердің құрамына кіреді.
- Биотин. Азот алмасуына қатысады, сонымен қатар қанықпаған май қышқылдарының синтезін катализдейді.
- Витамин РР(никотин қышқылы). Тотығу-тотықсыздану реакцияларына, ферменттердің түзілуіне және липидтер мен көмірсулардың алмасуына қатысады.
- Н витамині(параминобензой қышқылы). Бұл көптеген бактериялардың, соның ішінде адамның ішектерін мекендейтіндердің өсу факторы. Фолий қышқылы пара-аминобензой қышқылынан синтезделеді.
- Егіздер. Ол тотығу реакцияларына қатысатын кейбір ферменттердің құрамдас бөлігі болып табылады.
- Холин. Жасуша қабырғасының липидтер синтезінің реакцияларына қатысады. Ол аминқышқылдарының синтезіндегі метил тобының жеткізушісі болып табылады.
- Пурин және пиримидин негіздері(аденин, гуанин, ксантин, гипоксантин, цитозин, тимин және урацил). Заттар негізінен нуклеин қышқылдарының құрамдас бөлігі ретінде қажет.
- Амин қышқылдары. Бұл заттар жасуша белоктарының құрамдас бөлігі болып табылады.
Кейбір бактериялардың өсу факторларының қажеттілігі
Ауксотрофтарөмірін қамтамасыз ету үшін оларға сырттан химиялық заттарды жеткізу қажет. Мысалы, клостридиялар лецитин мен тирозинді синтездей алмайды. Стафилококктар лецитин мен аргининді қабылдауды қажет етеді. Стрептококктар май қышқылдарын - фосфолипидтердің компоненттерін қабылдауды қажет етеді. Коринебактериялар мен шигеллалар никотин қышқылын қабылдауды қажет етеді. Staphylococcus aureus, пневмококк және бруцеллалар В1 витаминін қабылдауды қажет етеді. Стрептококк және сіреспе таяқшалары – пантотен қышқылында.
Прототрофтарқажетті заттарды өз бетінше синтездейді.

Күріш. 24. Қоршаған ортаның әртүрлі жағдайлары бактериялар колонияларының өсуіне әртүрлі әсер етеді. Сол жақта - баяу кеңейетін шеңбер түріндегі тұрақты өсу. Оң жақта - «қашу» түрінде жылдам өсу.
Бактериялардың өсу факторларына қажеттілігін зерттеу ғалымдарға микробқа қарсы препараттарды, сарысуларды және вакциналарды өндіруде өте қажет үлкен микробтық массаны алуға мүмкіндік береді.
Бактериялар туралы толығырақ мақалалардан оқыңыз:
Бактериялардың көбеюі микробтар популяциясының санын көбейту механизмі болып табылады. Бактериялардың бөлінуі – көбеюдің негізгі жолы. Бөлінгеннен кейін бактериялар ересектердің мөлшеріне жетуі керек. Бактериялар қоршаған ортадан қоректік заттарды тез сіңіру арқылы көбейеді. Өсу үшін белгілі бір заттар (өсу факторлары) қажет, олардың кейбіреулері бактерия жасушасының өзі синтездейді, ал кейбіреулері сыртқы ортадан келеді.
Бактериялардың өсуі мен көбеюін зерттей отырып, ғалымдар микроорганизмдердің күнделікті өмірде және өндірісте қолданылуы тек қасиеттерімен ғана шектелетін пайдалы қасиеттерін үнемі ашып отырады.
Бактериялардың белсенділігі өсумен сипатталады- жасушаның құрылымдық және функционалдық құрамдас бөліктерінің қалыптасуы және бактериялық жасушаның өзінің көбеюі; сондай-ақ көбею- популяциядағы бактерия жасушаларының көбеюіне әкелетін өздігінен көбею.
бактериялар көбейедіекілік бөліну арқылы, сирек бүршіктену арқылы. Актиномицеттер саңырауқұлақтар сияқты споралар арқылы көбейе алады. Актиномицеттер тармақталушы бактериялар болып, жіп тәрізді жасушалардың бөлшектенуі арқылы көбейеді. Грам-оң бактериялар синтезделген бөліну бөлімдерінің жасушаға енуі арқылы, ал грам-теріс бактериялар тарылу арқылы бөлінеді, олардан екі бірдей жасуша түзілетін гантель тәрізді фигуралар пайда болады.
Жасушаның бөлінуі алдында болдыбактериялық хромосоманың жартылай консервативті түрге сәйкес репликациясы (екі тізбекті ДНҚ тізбегі ашылады және әрбір тізбек комплементарлы тізбекпен аяқталады), бактерия ядросының ДНҚ молекулаларының – нуклеоидтың екі еселенуіне әкеледі.
ДНҚ репликациясы үш кезеңде жүреді: инициация, ұзару немесе тізбектің өсуі және аяқталу.
Сұйық қоректік ортада бактериялардың көбеюі.Қоректік ортаның белгілі, өзгермейтін көлеміне егілген бактериялар көбейіп, қоректік заттарды тұтынады, бұл кейіннен қоректік ортаның таусылуына және бактериялардың өсуінің тоқтауына әкеледі. Мұндай жүйеде бактерияларды өсіру периодты культивация, ал дақылды периодты деп атайды. Егер жаңа қоректік ортаның үздіксіз берілуі және культура сұйықтығының бірдей көлемінің ағып кетуі арқылы өсіру жағдайлары сақталса, онда мұндай өсіру үздіксіз, ал культураны үздіксіз деп атайды.
Бактерияларды сұйық қоректік ортада өсіргенде түбіне жақын, диффузиялық немесе беткі (пленка түрінде) дақылдың өсуі байқалады. Сұйық қоректік ортада өсірілген бактериялардың мерзімді культурасының өсуі бірнеше фазаларға немесе кезеңдерге бөлінеді:
1. лаг фазасы;
2. логарифмдік өсу фазасы;
3. стационарлық өсу фазасы немесе максималды концентрация
бактериялар;
4. бактериялардың өлу фазасы.
Бұл фазаларды графикалық түрде бактериялардың көбею қисығының сегменттері ретінде бейнелеуге болады, ол тірі жасушалар санының логарифмінің олардың өсіру уақытына тәуелділігін көрсетеді.
Кешігу фазасы- бактерияларды себу мен көбеюдің басталуы арасындағы кезең. Арығу фазасының ұзақтығы орта есеппен 4-5 сағатты құрайды.Сонымен қатар бактериялар көлемі ұлғайып, бөлінуге дайындалады; нуклеин қышқылдарының, ақуыздың және басқа компоненттердің мөлшері артады.
Логарифмдік (көрсеткіштік) өсу фазасыбактериялардың қарқынды бөліну кезеңі болып табылады. Оның ұзақтығы шамамен 5-6 сағатты құрайды.Оңтайлы өсу жағдайында бактериялар әр 20-40 минут сайын бөліне алады. Бұл фазада бактериялар ең осал, бұл тез өсіп келе жатқан жасушаның метаболикалық компоненттерінің ақуыз синтезінің тежегіштеріне, нуклеин қышқылдарына және т.б. жоғары сезімталдығымен түсіндіріледі.
Содан кейін стационарлық өсу кезеңі келеді., бұл кезде өміршең жасушалардың саны өзгеріссіз қалады, максималды деңгейді құрайды (М-концентрация). Оның ұзақтығы сағаттармен көрсетіледі және бактериялардың түріне, олардың сипаттамаларына және өсіруге байланысты өзгереді.
Өлім фазасы бактериялардың көбею процесін аяқтайды, қоректік орта көздерінің таусылуы және ондағы бактериялардың зат алмасу өнімдерінің жинақталуы жағдайында бактериялардың өлуімен сипатталады. Оның ұзақтығы 10 сағаттан бірнеше аптаға дейін өзгереді. Бактериялардың өсу және көбею қарқындылығы көптеген факторларға байланысты, соның ішінде қоректік ортаның оңтайлы құрамына, тотығу-тотықсыздану потенциалына, рН, температура және т.б.
Бактериялардың тығыз қоректік ортада көбеюі.Тығыз қоректік ортада өсетін бактериялар бактериялық пигментке байланысты консистенциясы мен түсі әртүрлі, шеттері біркелкі немесе тегіс емес (S- және R-формалары) оқшауланған дөңгелектелген колониялар түзеді.
Суда еритін пигменттер қоректік ортаға диффузияланып, оны бояйды. Пигменттердің басқа тобы суда ерімейді, бірақ органикалық еріткіштерде ериді. Және, ең соңында, суда да, органикалық қосылыстарда да ерімейтін пигменттер бар.
Микроорганизмдер арасында ең көп таралған пигменттерге каротиндер, ксантофилдер және меланиндер жатады. Меланиндер – фенолды қосылыстардан синтезделген ерімейтін қара, қоңыр немесе қызыл пигменттер. Меланиндер каталаза, супероксидцисмутаза және пероксидазалармен бірге микроорганизмдерді улы оттегі асқын тотығы радикалдарының әсерінен қорғайды. Көптеген пигменттердің микробқа қарсы, антибиотикке ұқсас әсері бар.
Өсу қисығы белгілі бір орта жағдайында бактериялардың өсуі мен көбеюін сипаттайды. Өсу қисығы партия мәдениетін зерттеуден алынған.
мерзімдік мәдениетБұл қоректік заттардың жеткізілуінсіз қоршаған ортаның шектеулі көлемінде дамитын микроорганизмдердің популяциясы.
1 фаза – бастапқы – бактериялар өседі, бірақ көбеймейді
2 фаза – lg өсу фазасы – бактериялар қарқынды түрде көбейеді
3 фаза – стационарлық – көбею – өлімге тең
4 фаза – өлім – зат алмасу өнімдері жинақталады, қоректік орта таусылады, бактериялар өледі.
Сыртқы факторлар болуы мүмкін
- Бактериостатикалық әрекет- бактериялардың көбеюін және өсуін тежейді
- Бактерицидтік әрекет- бактерияларды өлтіреді
бактериялық ферменттер
- Ферменттер- химиялық реакцияларды катализдейтін арнайы белоктар. Ферменттер электрон тығыздықтарының қайта бөлінуін және субстрат молекуласының біршама деформациясын тудырады. Бұл молекулаішілік байланыстардың әлсіреуіне әкеледі, активтену энергиясы азаяды және реакция жылдамдайды.
Ферменттердің классификациясы -
- Катализденетін реакция түріне қарай – оксидоредуктазалар, лиазалар, трансферазалар, гидролазалар және т.б.
- Локализациясы бойынша – эндоферменттер – жасуша ішіндегі реакцияларды катализдейді. Экзоферменттер – бактерия жасушасынан бөлінетін, ыдырауды катализдейді
- Түзілудің генетикалық бақылауы – конститутивтік (барлық өмірлік циклде субстраттың болуы әсер етпейді), индукциялық – олар субстраттың болуына жауап ретінде қалыптасады.
- Субстрат бойынша – протеолитикалық – белоктарды, сахаролитикалық – көмірсуларды, липолитикалық – майларды ыдыратады.
Ферменттердің маңызы.
1. Ферменттердің синтезі анықталады, сондықтан ферментативті қасиеттерін анықтау организмдерді анықтауға қызмет етеді.
2. Бактериялардың ферменттері олардың патогенділігін анықтайды
3. Ферментативті қасиеттері микробиологиялық өнеркәсіпте қолданылады
Бактериялық ферменттерді анықтау
Протеазалар белоктарды аминқышқылдарына, мочевинаға, индолға, күкіртті сутекке, аммиакқа ыдыратады. Белоктары бар орталарда протеазалар осы өнімдерді бөліп алу арқылы анықталады. Желатинді қолданыңыз, ортаны сұйылту. Сүт сарысуында сұйылтуы бойынша және сүтте – мөлдірлігі бойынша. Казеин - ыдырайды, ақуыз коагуляцияланады. BCH-да индол газы мен күкіртті сутегінің бөлінуі үшін индикаторлық қағаздар арқылы анықталады
Көмірсуларды ыдырататын ферменттерді анықтау – сахаролитикалық. Бұл ферменттер көмірсуларды альдегидтерге, қышқылдарға, көмірқышқыл газына және Н2-ге ыдыратады. Оларды анықтау үшін MPB немесе MPA пайдаланыңыз, газ түзілу үшін қышқыл түзілу индикаторын + көмірсу + қалтқысын қосыңыз. Осы принцип бойынша Gis және Pestrel орталары құрылады. Егер қоршаған ортаның жарығы өзгерсе, газ бөлінсе, көмірсулар бөлінеді. Моносахаридтер қолданылады. Бұл принцип бойынша панельдер, таблеткалар, қағаз индикаторлық жүйелер, NIB - индикаторлық қағаздар жүйесі, энергетикалық түтік және ферментативті белсенділікті тіркеуге арналған құрылғылар жасалады.(Көмір қышқылы түзіледі => Ph бар индикаторлар қажет)
JSA – сары-тұзды агарда липолитикалық ферменттер – липазалар анықталады, оның құрамында сарысы бар, құрамында липидтер көп және липидтердің деструкциясы ортаның ағаруымен бірге жүреді.
Микроорганизмдерді өсіру.
Бұл қоректік ортада бактериялардың көп мөлшерін алу. Өсіру мақсаттары. үшін өсіру жүргізіледі
1. Микробиологиялық қасиеттерін зерттеу
2. Инфекцияларды диагностикалау үшін
3. Биологиялық өнімді алу үшін – бактериялардан немесе бактерияларды қолдану арқылы алынған.
Мұндай препараттар емдік, диагностикалық, профилактикалық болуы мүмкін. Бактерияларды өсіру жағдайлары
- Толық қоректік ортаның болуы.
- Оңтайлы температура
- Өсіру атмосферасы оттегі немесе оның болмауы.
- Өсіру уақыты - 18-48 сағаттан кейін көрінетін өсу, бірақ кейбіреулер - мысалы, туберкулез - 3-4 апта
- Жарық Кейбіреулер жарық болған кезде ғана өседі.
Аэробтарды өсіру әдістері
- Стационарлы – агардың бетінде
- Орташа аэрациямен терең өсіру әдісі. Аэрация қоршаған ортадағы оттегін еріту үшін жүргізіледі.
- Үздіксіз өсіру – ағынды қоректік орталарды пайдалану.
Микроорганизмдердің мәдени қасиеттері. Бұл қоректік ортада бактериялардың көбею ерекшеліктері.
Сұйық қоректік орталарда бактериялар ортаның лайлануын тудырады, тұнба түзе алады - түбіне жақын, париетальды және ортаның бетінде қабықша түзе алады. Колониялар тығыз қоректік орталарда түзіледі.
Колония- тығыз қоректік ортада бір түрдегі микроорганизмдердің оқшауланған жинақталуы. Оның белгілі бір өлшемі, беті, жиегі, пішіні, консистенциясы, құрылымы, түсі болады.
Колония түрлері
S-тегіс - пішіні дөңгелек, жиектері тегіс, беті тегіс.
R-колониялар – кедір-бұдыр, жиектері тегіс емес, беті жолақты
Колония SR 0 аралық - шеттері мен беті сәл тегіс емес.
Анаэробтарды өсіру ерекшеліктері. Анаэробтарды өсіру үшін оттегісіз жағдайлар жасалған. Бұл қол жеткізілді
- Қоректік ортаның регенерациясы – қоректік орта қайнап, еріген оттегі ортадан шығады.
- арнайы құрылғыларды – анаэростаттар мен эксикаторларды қолдану. Оларда оттегі не химиялық сіңіргіштермен жұтылады, не құрылғыдан сорылады.
- Ортаға тотықсыздандырғыш заттарды қосу – жеңіл және тез тотығатын заттар – көмірсулар, цистеин, паренхималық мүшелердің бөліктері, аскорбин қышқылы. Осы принцип бойынша анаэробтар үшін орта құрылды - Кит-Тарозци - анаэробтар ортасы. Оның құрамында BCH, көмірсулар және цистеин бар бауыр бөліктері бар.
- Тұқым себудің арнайы әдістері. Май астына себу, Вейон-Венян түтігіне себу, Фортнер бойынша егу. Аэробтар мен анаэробтар тостағанға қоныстанған - аэробтар оттегін сіңіріп, анаэробты орта алынады.
Таза дақылдарды оқшаулау.
таза мәдениет- сұйық немесе қатты қоректік ортада көп мөлшерде оқшауланған бір түрдегі микроорганизмдердің популяциясы.
Таңдау мақсаттары.
- Инфекциялардың диагностикасы. Таза дақылдарды бөліп алу бактериологиялық әдістің негізі болып табылады. Таза мәдениетті оқшаулауға және оны анықтауға негізделген. Идентификация - бұл түрдің анықтамасы.
- Биологиялық өнімдерді алу
- Бактериялардың биологиялық қасиеттерін зерттеу
- Санитарлық-гигиеналық зерттеулер
Аэробтардың таза дақылын бөліп алу кезеңдері.
- Қоспаны зерттеу – жағынды Грам бояуы.
- Қоспаны бөлу және колонияларды алу. Диссоциация жүргізіледі 1) Дрыгальский бойынша – агардың бетіндегі штрихтар. Цикл материалды алып, агарға егіңіз. Бірнеше шыныаяқтарға шпатель себу. 2) Сериялық сұйылту әдісі. 3) Кох – балқытылған агарда сериялық сұйылту әдісі.
- Колония жиілігін тексеру, жағынды, грамм бояу
- Таза культураны жинақтау үшін колониялардан агар көлбеуіне материалды субкультуралау. Таңдалған таза дақыл қасиеттері бойынша анықталады – морфологиялық, тинкториалды, культуралық, ферментативті және т.б.
Анаэробтардың таза дақылын бөліп алу
1. Анаэробтардың жинақталуы. Қоспа Kittarocy ортасына егіледі және 80 градус температурада 10 минут бойы қызады. Спора түзетін анаэробтар сақталады, ал басқалары – вегетативті формалары өледі. Содан кейін қоректік орта өңделеді, споралар өніп, жиналады
2. Цейслер бойынша колонияларды алу, анаэробты колонияларды агардың бетінде Анаэростатта, Вайнберг бойынша колонияларды Вейон-Вигнал түтіктерінде алады.
3. Колониялардың жиілігін тексеру – жағынды, Грам әдісімен бояу
4. Киттароциттік қоректік ортадағы колонияларды қайта егу, анаэробтармен жинақтау, таза дақыл.
5. Анаэробтың түрін анықтау, анықтау.
Таза мәдениеттерді оқшаулаудың басқа жолдары.
- Оңтайлы температураларды қолдануға болады
- Қоспаны 80 градуста 10 минут қыздырған кезде спораларды оқшаулау
- Шұңқырлау құбылысын қолдану – егіс алқабынан тыс таралу.
- Шукевич әдісі - сусымалы өсетін микроорганизмдердің таза дақылын бөліп алу.
- Бактериялардың сүзгіштігі – белгілі мөлшерде споралары бар сүзгілерден өту қабілеті. Қоспаны ультракүлгін сәулелермен, ультрадыбыстық, антисарысулармен өңдеу, осы факторларға төзімді микроорганизмдердің таза культурасын алу.
- Қоспаның электрофорезі арқылы. Белгілі бір заряды бар организмдер анодта немесе катодта жиналады.
- Микроманипуляторды қолданыңыз. Микроскоппен клетканы алып, бір микроб жасушасының төлін – клонды – таза дақылды алады. Элективті қоректік орталарды қолдану.
- Қоректік орталарға өт, тиурит тұздары, натрий хлориді, антибиотиктер қосылып, төзімді микроорганизмдердің таза дақылы бөлініп алынады.
- Сіз дифференциалды диагностикалық орталарды пайдалана аласыз.
- Сіз биологиялық әдісті пайдалана аласыз. Ақ тышқандар құрсақішілік бактериялар қоспасымен жұқтырылады және тропизмге байланысты бактериялар белгілі бір органға жиналады.
бактериялық пигменттер.
Бұл бактерия жасушасы бөлетін бояғыштар, олардың синтезі генетикалық түрде анықталады. Химиялық құрылымы бойынша пигменттер каротиноидтар – қызыл-сары, пирролдар – жасыл, фенозинді бояғыштар – көк-жасыл және меланин – қара ферменттер болуы мүмкін.
Сары – алтын түсті стафилококк, көк-жасыл – Pseudomonas aeruginosa
Пигменттер болып бөлінеді
- Ерімейтін пигменттер – тек колонияларды бояйды
- Еритін пигменттер – спирттерде, суда ериді
Пигменттер, әдетте, ауа микрофлорасында болатын бактерияларда, антибиотиктерге төзімді микроорганизмдерде түзіледі, өйткені. олар екіншілік метаболиттер және пигменттер көбінесе жарықтың қатысуымен түзіледі.
Пигменттердің қызметі
- Пигменттер зат алмасуға қатысады
- Антибиотиктерге төзімділікті арттыру
- Фотототықтыруға сезімтал аймақтарды қорғау арқылы ультракүлгінге төзімділікті арттырады
Л-бактериялардың формалары.
1935 жылы ашылған Бұл жасуша қабырғасы жоқ, бірақ өсу және көбею қабілетін сақтайтын микроорганизмдер. L формалары гетеротрофтар мен саңырауқұлақтардың көпшілігінде түзіледі. L түрленуін индукциялайтын факторлар -
1. Антибиотиктер
2. Амин қышқылдары – глицин, метионин, лейцин және басқалары.
3. Ферменттер – лизоцим.
4. Макроорганизмнің факторлары – макроденелер, комплимент
Бұл факторлар жасуша қабырғасын бұзады немесе жасуша геномына әсер етеді және жасуша қабырғасының компоненттерінің синтезі жүрмейді.
ҚасиеттерЛпішіндер.
- L пішіндері өзгермелі орта жағдайында бактериялардың тіршілігін қамтамасыз етеді.
- Бактериялардың кейбір түрлерінде морфологиялық ұқсас. Олар полиморфты – сфералық, грамтеріс. Олар А типті колонияларды – ортаның бетінде ұсақ колонияларды және В типті колонияларды – күңгірт орталық пен көтерілген жиектерді құрайды, колониялар қоректік ортаға өседі.
- Анаэробтар немесе микроаэрофильдер
- L-формаларының көбею жолдары көп – екілік бөліну, бүршіктену, бөлшектену, аралас.
- Олардың вируленттілігі төмендеген, адгезиясы жоқ, антигендік қасиеттері өзгерген.
- Олар кері қайтаруға қабілетті - бастапқы бактериялық пішініне оралу
Және емдеу қиын инфекцияларды тудырады.
Бұл L - формаларының антибиотиктерге төзімді болуына және олардың макроорганизмнің қорғаныш факторларына, антиденелерге, фагоцитозға, комплиментке төзімді болуына байланысты.
NFB бактерияларының өсірілмеген түрлері
Бұл метаболикалық белсенділікке ие, бірақ қоректік орталарда өспейтін бактериялар, көптеген микроорганизмдерде қолайсыз жағдайларға ұшыраған кезде дақылданбайтын түрге өтуін байқауға болады. Бұл ауысу генетикалық түрде бақыланады. Өту факторлардың әсерінен жүзеге асады
- Температура, әсіресе төмен
- Тұз концентрациясы
- Қоршаған ортаның аэрациясы
- Қоректік заттардың мөлшері
Өңделмеген формалардың құндылығы. Бұл пішінде олар эпидемиялар арасында сыртқы ортада сақталады және олар макроорганизмге енсе, оларды рекультивациялауға болады - қалпына келтіруге болады - бұл табиғи ошақты аурулардың болуын түсіндіреді.
Сәйкестендіру -
1. Тікелей жасушалар саны
2. ДНҚ белсенділігін анықтау
3. Генетикалық зерттеу әдістері.
Бактериялардың өсуі және көбеюі туралы түсініктер
Микробиологиялық диагностика, микроорганизмдерді зерттеу үшін, ал биотехнологиялық мақсатта микроорганизмдер жасанды қоректік орталарда өсіріледі.
астында бактериялардың көбеюібарлық жасушалық компоненттер мен құрылымдардың үйлестірілген көбеюі нәтижесінде популяциядағы олардың санын өзгертпестен жасушалардың массасының өсуін түсіну. Микроорганизмдер популяциясындағы жасушалар санының артуы терминмен белгіленеді «көбейту».Ол генерация уақытымен (жасушалар саны екі еселенетін уақыт аралығы) және бактериялардың концентрациясы (1 мл-дегі жасушалар саны) сияқты ұғыммен сипатталады.
Эукариоттардың митоздық бөліну циклінен айырмашылығы, прокариоттардың (бактериялардың) көпшілігінің көбеюі екілік, ал актиномицеттер бүршіктену арқылы жүреді. Сонымен қатар, барлық прокариоттар гаплоидты күйде болады, өйткені ДНҚ молекуласы жасушада сингулярлы түрде ұсынылған.
Бактериялық популяция
Бактериялардың көбею процесін зерттегенде, бактериялардың әрқашан көп немесе аз саны популяция түрінде болатынын және дамуын ескеру қажет. бактериялық популяциясұйық қоректік ортада партиялық культурада тұйық жүйе ретінде қарастыруға болады. Бұл процесте 4 кезең бар:
- 1-ші бастапқы,немесе кешігу фазасы,немесе тежелу фазасы,- жасушаның қарқынды өсуінің басталуымен сипатталады, бірақ олардың бөліну жылдамдығы төмен болып қалады;
- 2 - логарифмдік,немесе журнал фазасы,немесе экспоненциалды фаза,- жасушаның бөлінуінің тұрақты максималды жылдамдығымен және популяциядағы жасушалар санының айтарлықтай өсуімен сипатталады;
- 3-ші - стационарлық фаза- популяциядағы жасушалар санының көбеюін тоқтатқанда пайда болады. Бұл жаңадан пайда болған және өлетін жасушалардың саны арасында тепе-теңдік бар екеніне байланысты. Стационарлық фазадағы қоректік ортаның көлемі бірлігіне келетін популяциядағы тірі бактерия жасушаларының санын М-концентрация деп атайды. Бұл көрсеткіш бактериялардың әрбір түріне тән белгі болып табылады;
- 4 - өлу фазасы (логарифмдік өлім)- популяциядағы өлі жасушалар санының басым болуымен және популяциядағы өміршең жасушалар санының үдемелі төмендеуімен сипатталады. Микроорганизмдер популяциясының санының өсуінің (көбеюінің) тоқтауы қоректік ортаның таусылуына және/немесе ондағы микроб жасушаларының зат алмасу өнімдерінің жиналуына байланысты болады. Сондықтан метаболизм өнімдерін жою және/немесе қоректік ортаны ауыстыру, микроб популяциясының стационарлық фазадан өлу фазасына өтуін реттеу арқылы белгілі бір деңгейде динамикалық тепе-теңдікті жоюға ұмтылатын ашық биологиялық жүйені құруға болады. халықтың дамуы.
Бұл микроорганизмдердің көбею процесі деп аталады ағындық мәдениет (үздіксіз мәдениет).Үздіксіз культурада өсу арнайы құрылғыларда (химостаттар мен турбидистаттар) ағынды өсіру кезінде бактериялардың үлкен массасын алуға мүмкіндік береді және вакциналар өндірісінде, сонымен қатар микроорганизмдер өндіретін әртүрлі биологиялық белсенді заттарды алу үшін биотехнологияда қолданылады.
Жасушаның бөліну цикліндегі метаболикалық процестерді зерттеу үшін де қолдануға болады синхронды дақылдар- популяциясының барлық өкілдері циклдің бір фазасында болатын бактериялардың осындай дақылдары. Бұған арнайы өсіру техникасы арқылы қол жеткізіледі.
Дегенмен, бірнеше бір уақытта бөлінуден кейін синхрондалған жасуша суспензиясы біртіндеп асинхронды бөлінуге қайта ауысады, осылайша ұяшықтар саны одан әрі сатылап емес, үздіксіз өседі.
Колониялар
Тығыз қоректік орталарда өсіргенде бактериялар түзіледі колониялар- жай көзге көрінетін, көбінесе бір жасушаның ұрпағы болып табылатын бір түрдегі бактериялардың жиналуы.
Түрлі бактериялардың колониялары әртүрлі:
- пішін;
- мөлшері;
- ашықтық;
- түс;
- биіктігі;
- бетінің және шеттерінің сипаты;
- консистенциясы.
Колониялардың табиғаты бактериялардың таксономиялық ерекшеліктерінің бірі болып табылады.


 Кейіпкерлердің ит жүрегінің қысқаша сипаттамасы")


